| REPRINTED FROM |
| Proceedings of the International Symposium on |
| AFRICAN VERTEBRATES |
| SYSTEMATICS, PHYLOGENY AND EVOLUTIONARY ECOLOGY |
| KARL-L. SCHUCHMANN, Editor |
| ZOOLOGISCHES FORSCHUNGSINSTITUT UND MUSEUM ALEXANDER KOENIG, BONN |
| THE LIZARD GENUS TETRADACTYLUS: A MODEL CASE OF AN EVOLUTIONARY PROCESS |
| By HARTWIG A. E. BERGER-DELL'MOUR State Museum, P. O. Box 1203, Windhoek 9000, SWA/Namibia * |
| Introduction |
| Ever since the concept of evolution was established, it has been one of the major objectives of zoology to formulate models on how evolution actually takes place. The processes that are involved take too long to be monitored even by a biologist with a long life-span. Some results can be obtained by working with low-key organisms such as bacteria, but they may only be generalized with restriction, since these organisms differ too much from the higher metazoa, evolution theory is usually focused on. |
| A major problem is the lack of "missing links". Paleontological evidence is scarce and even in well represented cases (like the transitional series of horse-limbs) is restricted to bones. |
| An investigation of the missing links as living organisms will not be possible before time-travelling machines have been invented. For those, however, who don't want to wait so long there is still the possibility to work on a group of closely related animals that show a gradation of forms. If the conditions under which the transition occurred remained constant and if monophyletic origin ensures that the genetic set-up of all recent species was initially the same, one might duly assume that today's intermediate forms resemble the missing links between ancestor and the most advanced living form. |
| The case of the grass-lizard Tetradactylus from southern Africa was considered to fit into this concept. In an extensive monograph (Berger-Dell'mour 1983) the functional aspects of transition from a running tetrapod lizard to a legless snake-like "Grasschleiche" were examined. The paper at hand contains a brief summary of the results and discusses their indications. |
| For detailed descriptions of anatomy and analyses of locomotion, BERGER-DELL'MOUR (1983) should be consulted. |
| Material and Methods |
| 31 specimens (15 + 7 + 9) of Tetradactylus s. seps (Linnaeus), Tetradactylus t. tetradactylus (Lacepede) and Tetradactylus a. africanus (Gray) were collected in dense grassland along the south and east coasts of South Africa. Live lizards were filmed from above at 45 frames/sec while moving over different types of surface ranging from smooth glass to a layer of dry grass. |
| The anatomy of the locomotory organs was examined and compared, and a biometrical analysis of body-shape and of sceletal elements that have obviously changed during the course of evolution was done. |
| Results |
| The first step |
| The main conclusion to be drawn was that transition is primarily functional and only secondarily morphological. This is proven by the most "primitive" species of the three, Tetradactylus seps, which hardly shows any difference in body shape compared to a "normal" lizard but is able to fold down both pairs of limbs in dense grass and move like a snake ("Schl�ngeln" Wiedemann 1932; "serpentine movement" Gray & Lissmann 1946; "lateral undulation" Gans 1962; "funktionelles St�tzschl�ngeln" Berger-Dell'mour 1983). |
| The selective advantage of this ability was demonstrated to me, as the actual "predator" on the spot, in a little patch of grassland on the Cape Peninsula, where Tetradactylus seps occurs together with a skink of about the same size, Mabuya homalocephala. After a short early summer's rainstorm the lizards were warming up in the top layer of the grass patch. On being approached, both kinds of lizard would dive into the dense grass. The skink, however, could be heard rustling on the ground, and occasionally, one would even see blades of grass moving at the top. |
| On the other hand, all one would hear or see of Tetradactylus seps was a quick movement when it took fright. It was clear that it moved on in the dense grass for quite some distance, but for the human observer it was impossible to determine in which direction and for how far. A bird of prey or a mongoose would definitely have more chance to catch a Mabuya than a Tetradactylus in that area. The possibility of approaching prey unapprehendedly should also be of advantage to the lizard. |
 |
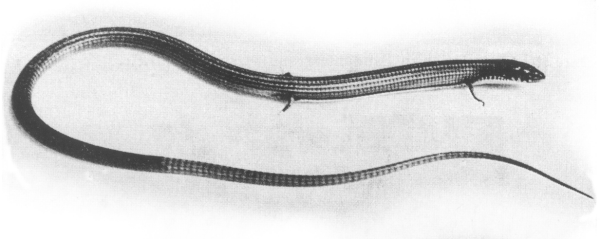
| Fig. 1: Tetradactylus s. seps. |
 |
| Fig. 2: Tetradactylus t. tetradactylus |
 |
| Fig. 3: Tetradactylus a. africanus |
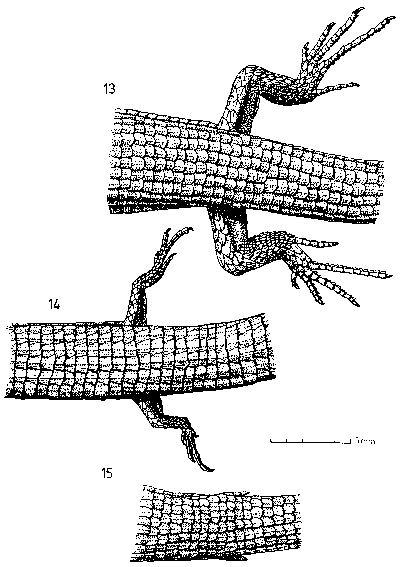 |
| Fig. 4: Pelvic region from above; (13) T. seps, (14) T. tetradactylus, (15) T. africanus. |
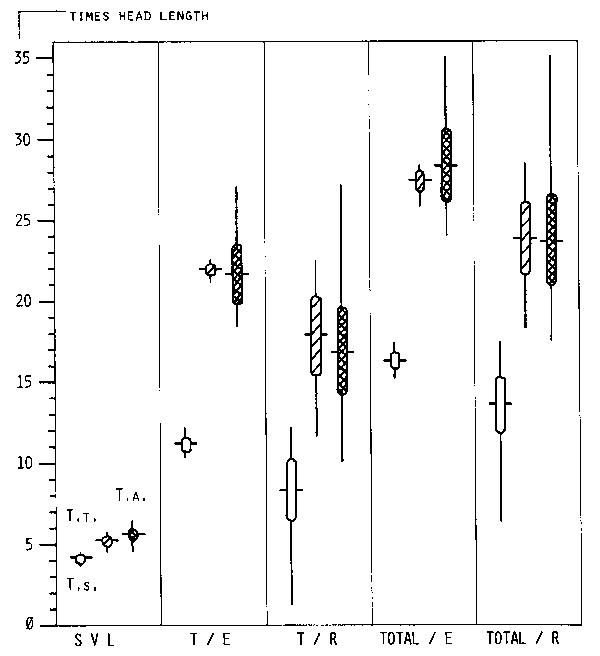 |
| Fig. 5: Body proportions in the three species. (SVL: Snout-vent length; T/E: Tail length of specimens with entire tail; T/R: Tail length of specimens with broken or regenerated tail; TOTAL/E: Total length of specimens with entire tail; TOTAL/R: Total length of specimens with regenerated/broken tail.) Measures standardized by head length. Vertical line: range; horizontal line: mean. T. t. and T. a. have almost the same range and means. Both together are clearly separated from T. s. |
| Priority to more advantageous changes |
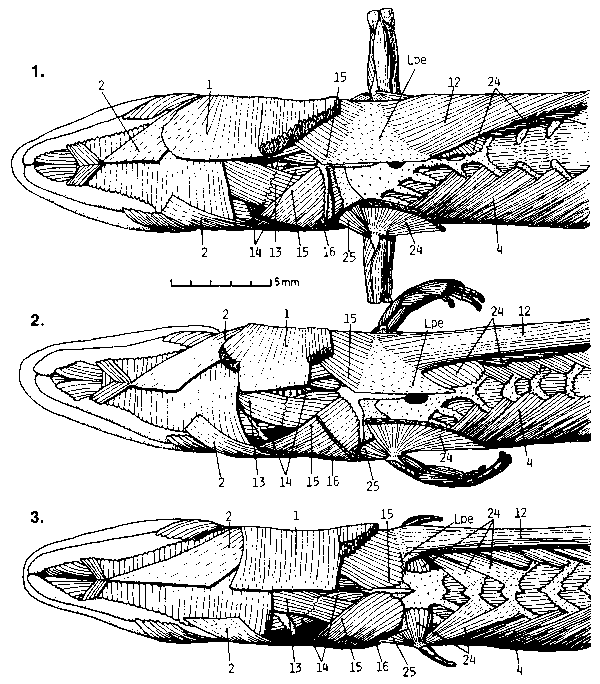
| Another observation was that all structural changes, whether constructive (like the increase of body segments) or destructive (rudimentation of limbs and their components) show a pattern of high selectivity. The size of the limbs changes early in evolution. The intermediate species, T. tetradactylus, thus possesses miniaturized extremities. Limb structure, however, remains largely unaltered. The species still possesses most of the bones of a "normal" lizard (see Fig. 7) and a complete set of muscles. |
| The general body shape of a "Schleiche" has already been attained in T. tetradactylus. That is, the mean index of body length over head length (as a standard) of T. africanus is only slightly greater than that of T. tetradactylus: The swiftness for moving through the grass like a snake should be much improved if the overall length of body with a constant diameter (see Gans 1975) that is used in locomotion increases. Similarly, miniaturized limbs facilitate gliding past stems of grass. |
| The final rudimentation of limbs and interior locomotory organs should only be advantageous in the sense of "saving building material": Lizards might much more often survive by escaping from a predator through their swiftness than by enduring their embryonic development because of a few more milligram of unused yolk at their disposal. |
| Variation, asymmetry and the importance of ontogenetic processes. |
| There is no typical body shape in one species as the typological concept would make us believe. One may only refer to a certain breadth of variation for every organ that is involved in the transitional process (see Figs. 7 to 11). |
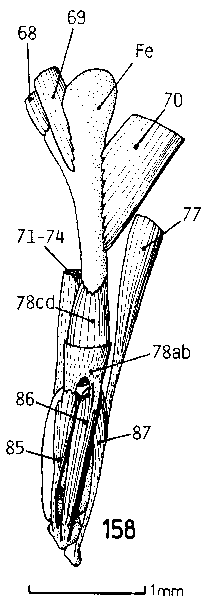 |
| Fig. 6: Left hindleg, dorsal, of T. africanus. Muscles 85 & 87 are functionless on the minute styloid limb rudiment. No moving of the small tarsal or metatarsal bones (0,4 and 0,2 mm long) would be possible, since skin and scales around them are too rigid. |
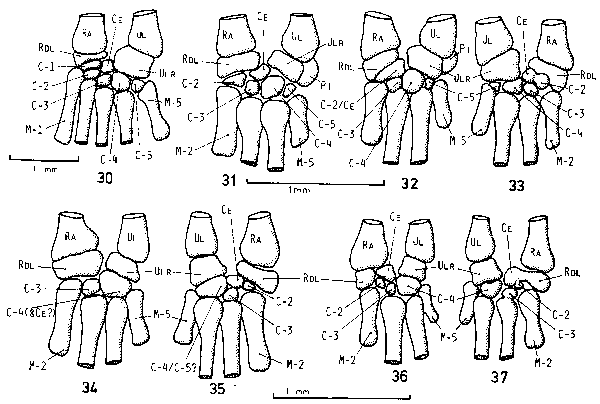 |
| Fig. 7: Base of hand, left to right, dorsal. 30: T. seps; all bones of a normal lizard are present. 31 - 37: T. tetradactylus; almost all of the bones may be present (31) but the extremity has considerably been reduced in size. Note fusion of bones and the great extent of variation, even in one individual (32 & 33, 34 & 35, 36 & 37 are the hands of one individual each). |
 |
| Fig. 8: T. africanus; base of hand, dorsal. Note variation of rudimentation within one population. |
 |
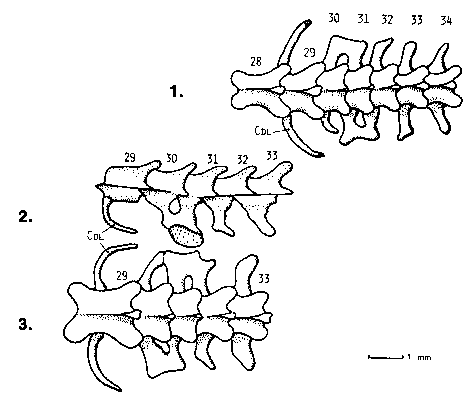
| Fig. 9: Diagram of precaudal vertebrae, ribs and two muscles, indicating the extent of Variation within and between species. (Cce: Cervical ribs; Cdl: Dorsolumbal ribs; Cpa: Parasternal ribs; Cst: Sternal ribs; Cxi: Xiphisternal ribs; FREE RIBS: Ribs with lose ends, attached to skin by muscles only; M.4: Musculus obliquus externus, pars superficialis; M. 10: Musculus intercostalis scalaris; SKIN-RIBS: Ribs attached to the inside of the ventral skin by their tips; Vca: Caudal vertebrae; Vce: Neck vertebrae; Vdl: Dorsolumbal vertebrae; Vdo: Dorsal vertebrae; Vsa: Sacral vertebrae; Vth: Thoracic vertebrae.) |
| Variation even occurs between the two sides of the body (in which case it is called asymmetry). This lays evidence that the change from one structure to another might be a question of embryological development rather than of well-defined genetic changes: If, for example, we accept that elongation of the body (increase of presacral vertebrae) is caused by a shift of the pelvis-inducing limb bud rather than by a series of coordinate mutations that change the genetic code for the construction of a sacral somite to that of a presacral and of a caudal to a sacral one, including harmonic changes of all the organ systems, we will face fewer problems in understanding the large number of morphologically different individuals in one population (see Fig. 9). It also gives a more plausible explanation for asymmetrical shape of pelvic or sternal bones (Figs. 9 and 10). |
| Van Bemmelen's theory of the splitting of embryonic somites |
| Van Bemmelen (1952), in a theoretical paper based on findings of earlier authors, suggested that the position of pectoral and pelvic girdles is genetically fixed and that in cases where the number of presacral vertebrae is increased, this is due to a splitting of embryonic somites. He proposed that in most snakes and snake-like lizards the number of vertrebrae between neck and sacrum would be twice or n-times the original number in reptiles (which he claims to be 24). An increased number of vertebrae in neck and tail would be due to "other factors". |
| This theory has been shown to be not valid. Furthermore, asymmetrical organization of the thoracic ribs and of the sacrum proves that the position of the girdles is all but fixed. |
 |
| Fig. 10: T. seps, sacral region. 1. & 3. dorsal; 2. from left. Note asymmetrical structure of sacrum. Drawings 2. & 3. refer to the same individual. |
| "Trends" |
| Some authors have assumed that evolutionary processes like the transition from lizard to "Schleiche" take place in a well-coordinated, harmonic and teleonomic way. Others have thought that there is at least a genetic linkage between elongation of the body and reduction of limbs (Boeker 1935; Raynaud 1972, 1974, 1976 a & b). |
| In the case of Tetradactylus, there is no evidence of such trends. The one specimen of T.seps with maximally reduced arms and legs had the most "primitive" snout-vent length and a low number of 29 presacral vertebrae (most specimens have 30) which would be impossible if there was a common genetic denominator for reduction of limbs and increase of segments. |
| The question of teleonomic evolution was tried to be assessed by judging 25 features of each specimen, placing its position within the species' limits of variation. It turned out that almost every animal was very primitive in some aspects and highly advanced in some others. There was no individual that was generally "behind" or "ahead" of the others (the means were usually close to the mean of the species as a whole). This would not be so if there were "forces" that actively change the genome towards a more advantageous structure. |
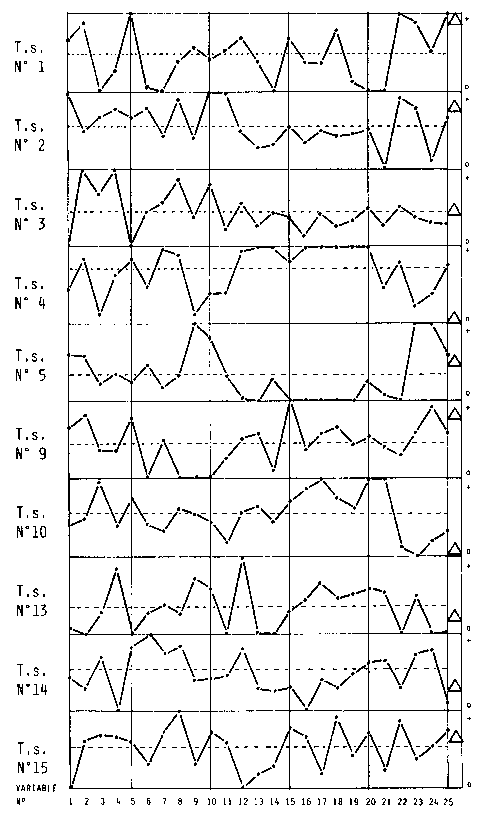 |
| Fig. 11: Variation of 25 features that change characteristically between the species in ten individuals of T.seps (1-6: Thorax; 7-12: Shoulder; 13-15: Front leg; 16-18: Hind leg; 19-25: Pelvis). The bottom line of each block indicates the most "primitive" value of the sample (marked "o"), the top line shows the most "advanced" value (marked " + "). Each block shows the situation of one specimen. Every individual is primitive in some aspects and advanced in others. The triangle indicates the relative snout-vent length. For number of presacral vertebrae see Fig. 9. Note that in the specimen with lowest SVL of the sample (T.s. 4) arms and legs (features 13-18) show the highest degree of rudimentation. (Measures standardized against head length to avoid artificial "readings".) |
| Grass-gliding versus burrowing "Schleichen" |
| There seems to be a fundamental difference between grass frequenting and burrowing snake-like lizards which appears if the number of presacral vertebrae is plotted against snout-vent length (head length would have been better, but the data from Gasc 1979 that were used to demonstrate the situation in a burrowing skink, did not include this measure). |
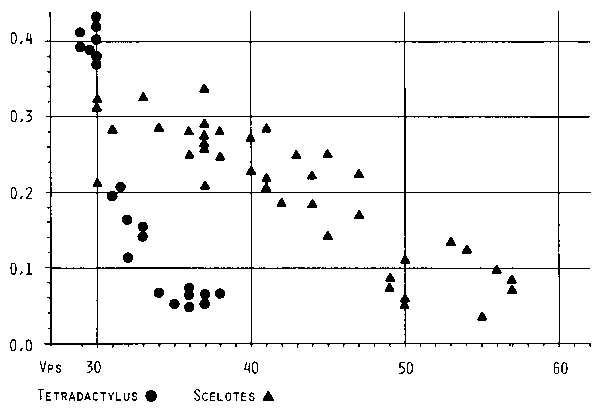 |
| Fig. 12: Relative length of hind leg (vertical axis, in proportion to snout-vent length) plotted against number of presacral vertebrae (horizontal axis) in the grass-gliding genus Tetradactylus and the burrowing genus Scelotes. (Data for Scelotes extracted from Gasc 1979.) |
| Burrowing skinks basically enlarge their presacral region, grass lizards mainly the tail. There seems to be no apparent reason for this, except that it might be much "faster" in terms of evolutionary processes to increase tail length. For digging underground (which makes "concertina movement" necessary) the tail part of the body with its specific musculature might be useless. "Grasschleichen" that have the same relative body length as burrowing "W�hlschleichen", might thus be much less advanced in general terms of evolutionary development. The relatively long limbs of Tetradactylus seps compared with Scelotes having the same number of presacral vertebrae confirm this (see Fig. 12). |
| The Musculus rectus lateralis issue |
| A particular muscle (M. rectus lateralis) was considered by Camp 1923 to be of major significance. Camp divided the whole group Lacertilia into two sections, the "Autarchoglossa", possessing this muscle and the "Ascalabota" without it. Only Autarchoglossa would be preadapted to become snake-like, because the muscle would enable them to perform undulatory locomotion. His classification proved to be artificial when the Pygopodids were found to be closely related to the geckos. Furthermore, in the genus Tetradactylus, the M. rectus lateralis is more powerful in T.seps than in T.africanus, thus showing Camp's theory on the origin of snake-like lizards to be invalid (see Moody 1983). |
 |
| Fig. 13: Outer layers of muscles of throat and thorax. The Musculus rectus abdominis, pars lateralis (12) is weaker in the more snake-like species. |
| Locomotion analysis |
| In every species the following statement is valid: The smoother the surface, the more "primitive" (tetrapod-like) the locomotory movements are. |
| On a smooth glass plate, T.seps uses its legs for running in the typical "cross-walk" fashion of a tetrapod. This pattern of moving right foreleg and left hindleg simultaneously and alternately to left foreleg and right hindleg also shows in T.tetradactylus. Body undulations, though, do not correspond to the movements of legs, and alterations in the cross-walk pattern occur. |
| Both species show a series of transitional stages of locomotion that range from "swimming movements" (both arms are moved simultaneously instead of alternately) and pressing one hindleg against the body on the outside of the curvature to "Quasi-Schl�ngeln" (both hindlegs are continuously pressed to the body; the forearms move, obviously groping for "contact-points") and finally true snake-like movement "Funktionelles St�tzschl�ngeln", in dense vegetation. |
| T.tetradactylus is not able to move forward on glass, although it can walk on a smooth wooden board, where the ratio of static friction of the body against that of the legs is obviously better. T.africanus is not able to move forward at all if there are no lateral contact points available for "Schl�ngeln". |
 |
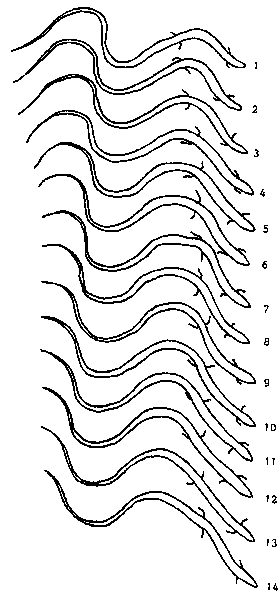 |
| Fig. 14: One walking cycle in T.seps. Stride and body curvature correspond. |
| Conclusion |
| For reasons given above, the transitional stages within the genus Tetradactylus seem to be a good example of evolutionary change. |
| The selective advantage of an alteration and its manifestation are proportional, thus resulting in a seemingly well-balanced course of evolution. This should, however, not lead to teleonomic interpretation. |
| No evidence of "guiding forces" was found, and a genetic link between constructive and destructive evolution of organs could not be confirmed. |
| The first step in the transition from a lizard to a snake-like organism is obviously a change in locomotion (i. e. behaviour) rather than either reduction of limbs or elongation of the body "in advance". The difficulty in explaining morphological "preadaptations" seemingly having no initial selective advantage is not encountered in this evolutionary model. |
| An apparent "orthogenetic trend" should not lead to the conclusion that "Orthogenesis" exists as a propulsive force. There is no reason to believe that T.tetradactylus, set free on an island with no reptiles, but many ecological niches for running lizards vacant, would not split into a few ecotypes, some of them again having fewer vertebrae and longer legs (presumably with only four digits, though). The reason why no Tetradactylus "goes back into the open areas" rather seems to lie in its specialization to a certain biotope and the abundance of competitors which might just be that little bit "fitter" outside this biotope. |
| Models of adaptive radiation |
| Four models of how speciation in a group of grass-gliders might have taken place are shown in Fig. 16. |
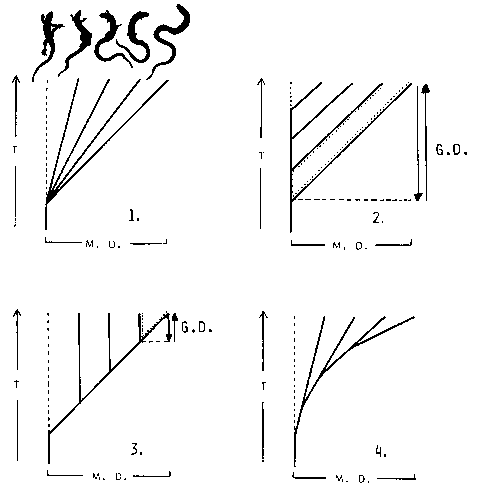 |
| Fig. 16: Four models of how adaptive radiation in a group of "Schleichen" might take place. (G. D.: Genetic distance; M. D.: Morphologic distance [degree of dissimilarity]; T: Time.) |
| 1. One possibility would be that all forms separated from their common ancestor at the same time, and have since evolved at different speed. |
| 2. From a lizard-like type that did not change its morphology, successive "off-splittings" of populations towards the new morpho-eco-type might have occurred. From the moment of separation onwards, the speed of structural change could have been the same for every group. |
| 3. The whole group developed towards the new form. In certain areas, the ecological "target niche" (Gans 1975) was found occupied by another, more advanced grass-glider, and evolution stopped, leaving the population in "equilibrium with a current set of environmental conditions" (Gans 1975). |
| 4. Finally, there might have been a repeated splitting-off of marginal populations which then developed at speeds different from the parental population. |
| Of course, any combination between the models would have been possible. |
| An investigation of the true "genetic distance" by DNA-DNA hybridization (Sibley & Ahlquist 1983) should enable us to recognize which course was actually taken. |
| For such a study, four more species of Tetradactylus, that have to be placed between T.tetradactylus and T.africanus, according to the degree of limb-reduction, and two "beyond" T. africanus would be available. (In the 1983 study, it was too difficult to obtain specimens of these scarce species.) |
| If it turns out that genetic distances are smallest between the most "advanced" forms, there would be some uncertainty whether model 3 or 4 is applicable. It would have to be decided against model 3, which seems to be quite unrealistic: With all three species occurring in strikingly similar kinds of habitat, the "equilibrium" theory does not seem to apply. |
| Besides, the only species that lives together with a grass-gliding competitor (Chamaesaura anguina) in most of its range is Tetradactylusafricanus - the most snake-like of the three. Furthermore, on the Cape Peninsula, where T.seps and T.tetradactylus occur sympatrically (not syntopic), the former inhabits denser grass patches in the lowlands and is generally more widely distributed. It cannot be seen as "having been driven into refuge biotopes by the superior competitor". |
| Summary |
| 31 individuals of three species of the genus Tetradactylus Merrem were collected in 1978 in South-Africa. From T.seps through T.tetradactylus to T.africanus, they show a transition from normal, lizard-shaped reptile to snake-like "Schleiche". Anatomical investigation of the locomotory organs, biometrical studies and analysis of locomotion, accompanied by field observations, revealed that the first stage of transition is behavioral rather than morphological: Tetradactylus seps, although lizard-like in shape and capable of tetrapod locomotion on a smooth surface, will fold its legs and switch over to snake-like undulation whenever it quickly moves through dense grass thickets, its natural habitat. |
| The overall body-shape is attained early in the evolutionary process, while final rudimentation of limbs occurs considerably later. The pattern of morphological changes thus follows a strategy by which priority is given to more advantageous alterations. |
| "System-immanent" propulsive forces that would produce a harmonically coordinated or even teleonomic course of evolution could not be traced and, contrary to some earlier publications, no evidence of genetic linkage between rudimentary and constructive evolution was found. |
| References |
| Berger-Dell'mour, H. (1983): Der �bergang von Echse zu Schleiche in der Gattung Tetradactylus, Merrem. - Zool. Jb. Anat. 110: 1-152. |
| Boeker, H. (1935): Einf�hrung in die vergleichende biologische Anatomie der Wirbeltiere. I. Band. - Verlag Gustav Fischer, Jena. |
| Gans, C. (1962): Terrestrial locomotion without limbs. - Amer. Zool. 2: 167-182. |
| - (1975): Tetrapod limblessness: evolution and functional corollaries. - Amer. Zool. 15: 455-467. |
| Gasc, J. P. (1979): �tude des modalit�s de r�duction des membres chez un Squamate serpentiforme: Scelotes, Scincid� afro-malgache. - Ann. Sci. Nat. Zool. Paris, 13e Ser., 1: 99-132. |
| Gray, J. (1946): The mechanism of locomotion in snakes. - Journ. Exp. Biol. 23: 101-120. |
| Moody, S. (1983): The Rectus abdominis muscle complex of the Lacertilia: Terminology, homology, and assumed presence in primitive Iguanian Lizards. - In: Advances in Herpetology and Evolutionary Biology. Essays in Honour of Ernest E. Williams. A. G. J. Rhodin & K. Miyata (eds.), pp. 195-212. |
| Raynaud, A. (1972): Morphogen�se des membres rudimentaires chez les Reptiles: un probl�me d'embryologie et d'�volution. Bull. Soc. Zool. France 97: 1171-1174. |
| - (1974): Donn�es embryologiques sur la rudimentation des membres chez les Reptiles. - Bull. Soc. Hist. Nat., Toulouse, 110: 26-40. |
| - (1976 a): Les diff�rentes modalit�s de la rudimentation des membres chez les embryons des Reptiles serpentiformes. - Coll. Int. CNRS, Toulouse, No. 266, pp. 201-219. |
| - (1976 b): La rudimentation des membres des Reptiles au cours de l'ontogen�se. - Bull. Soc. Zool. France, Suppl. No. 101, pp. 1-3, 40-46. |
| Van Bemmelen, J. F. (1952): Lichaamsverlenging slang en hagedis. - Proc. Kon. Nederl. Akad. Wetensch., Ser. C, 55: 215-237. |
| Wiedemann, E. (1932): Zur Ortsbewegung der Schlangen und Schleichen. - Zool. Jb. Physiol. 50: 557-596. |
| * address not valid |
| Fig. 15: Three walking cycles in T.tetradactylus. Body curvature is not coordinated with stride. |
| Sibley, C. G., & J. E. Ahlquist (1983): Phylogeny and classification of birds based on the data of DNA-DNA hybridization. In: Current Ornithology, Vol. 1, R. F. Johnston (ed.), Plenum Publishing Corp. |
 |
 |