"I am suggesting here that organisms have a built-in capability of adapting to their environment".
(Lee Spetner,"Not By Chance!", 1998, sivu 200)
(Kiitos:HumppaaSaatanat)
Lee Spetner on informaatioteoreetikko, joka kirjassaan "Not by Chance!" on esitt�nyt, ett� "Satunnaiset mutaatiot eiv�t voi tuottaa evoluution vaatimia informatiivisia muutoksia, eli tuottaa uutta informaatiota." Kirjassaan, sivulla 210, h�n sen sijaan kertoo kannattavansa "eisattumanvaraista variaatiota", johon h�net on inspiroinut Talmud. Spetnerin omassa mallissa, jota h�n kutsuu nimell� "Ei-sattumanvaraisesta evoluutiohypoteesista"("Non Random Evolutionary Hypothesis"~"NREH"),mutaatiot siis ovat itse asiassa "enimm�kseen hy�dyllisi�", ja niiden ilmestyminen yli�lyn ohjelmoimia. Spetnerin mukaan mutaatiot eiv�t varsinaisesti "tuota informaatiota" vaan nostavat esiin "perim�n potentiaalista informaatiota" esimerkiksi her�ttelem�ll� "nukkuvia geenej�"(~Spetnerin "dormant genes").
Spetner ei kuitenkaan koko h�nen kirjassaan k�sittele "Evoluution suuntaajaa" eli luonnonvalintaa muuten kuin muodossa "se karsii informaatiota", ottamatta huomioon "mink�laista informaatiota". N�inollen esimerkiksi Dawkinsin esitt�m�t "luonnonvalinnan rajoittamat, ennustettavat evoluutiopolut" j��v�t k�sittelem�tt� ja evoluutio k�sitell��n pelk�st��n "perinteisist� perinteisimm�ll�" evoluutio -olkinukkena, "puhtaansa sattumana". Parhaimmillaankin Spetner siis todistaa, ett� eli�kunta ei ole syntynyt kerralla sattumalta, mik� onkin aivan j�rkev��. Spetnerin n�kemys painottaa sit�, ett� mutaatiot ovat "eisatunnaisia", ja siksi hy�dyllisi� ja ett� "satunnaisuus ei informaatiota voisi tuottaa". Spetnerin suurin varsinainen ero "vallitsevaan uusdarwinismiin" onkin juuri se, ett� h�nen mallissaan valinta vaihtoehdoista on "a priori"(�lykk��n tahon koodaama "mikroevoluution potentiaali") ja evoluutiossa "a posteriori"(luonnonvalinta).
Todistaakseen n�kemyksens� oikeaksi, tulisi Spetnerin siis osoittaa
miksi se kun joku �lykk��n tahon ohjelmoimat s��nn�t valitsevat "vapausasteisista systeemeist� funktionaalisesti sopivat", lis�� informaatiota, mutta jos luonnonvalinta tekee saman, niin informaation m��r�n kasvua ei voi tapahtua. T�m� voisi tapahtua esimerkiksi esitt�m�ll� tavan, jolla "potentiaalinen informaatiom��r�" voitaisiin selv�sti esiinsaada.
Spetnerin Informaatiom��rittely
"all point mutations that have been studied on the molecular level turn out to reduce the amount of information"
(Lee Spetner, "Not By Chance!", 1998, sivu 138)
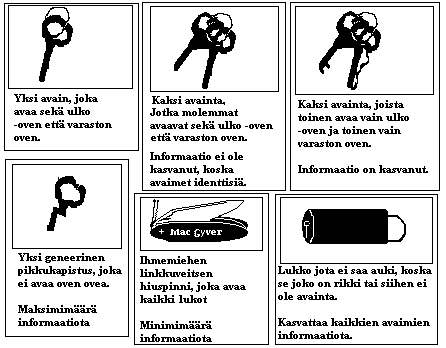
Spetner soveltaa informaatiomalliaan entsyymeihin ja n�iden suhteeseen olemassaoleviin substraatteihin ja n�iden lukum��r��n. Spetnerin informaatiolla on kaksi piirrett�:
1: "odotusmittaus" ("expectation" measure) : Erilaisten merkkijonojen kokonaisuudella on v�hemm�n informaatiota kuin identtisten merkkijonojen kokonaisuudella.
2: "osoitemittaus" ("addressing" measure of information") : Pitemm�ss� merkkijonossa on enemm�n informaatiota. Informaatio on "kyll� tai ei"~"kaikki tai ei mit��n" -tyyppist�.
Spetnerin informaation m��r� kasvaa sit� suuremmaksi mit� enemm�n on substraatteja joihin entsyymi ei vaikuta. Eniten informaatiota on "spesifisess� entsyymiss�", joka tehoaa ainoastaan yhteen substraattiin.
M��rittely ja mitattavuus
"When I say a variation is not random, I mean that the chance of it occurring has something to do with how the organism adapts to its environment"
(Lee Spetner, "Not By Chance!", 1998, sivu 175)
Spetnerin m��rittelyn pohjalta luonnossa ilmenev�n informaation mittaaminen on mahdotonta, koska se vaatisi sen, ett� tutkittavan entsyymin toiminta tulisi testata suhteessa joka ikiseen universumissa mahdolliseen substraattiin : Ja n�iden lukum��r�� saati rakennetta emme edes tied�, saati sitten saa yhteiseen tilaan entsyymin kanssa.
Spetnerin informaatiomallissa oletetaan,ett� geeni toimii "joko-tai" mekanismilla ilman v�livaiheita; t�m�n ns. geeniperheet osoittavat v��r�ksi : niiss� useat samankaltaiset geenit hoitavat erilaisia, joskus osittain limitt�isi�kin, toimintoja. (Lokki, Saura & Tigerstedt,"Evoluutio ja populaatiot.",1986) N�inollen toiminto voi muuttua ja kun kahdentuminen on mahdollista, voi kahdentuma + toisen muutos johtaa uuden informaation syntyyn "eritt�in mikroevolutiivisesti".
Spetnerin informaatiom��rittelyss� on my�skin joitain puutteita. Siin� ei esimerkiksi oteta lainkaan huomioon toimintalaatua. Jos entsyymi toimii mitenk��n on se informaatiota; Spetnerin informaatio on "joko tai", vaikka entsyymien toiminta ei t�ll�ist� ole, ja eli�n toiminnallisuuden kannalta on kuitenkin suunnatonta merkityst� sill� mill� teholla entsyymi toimii."Entsyymi - substraattiyhdistym� muodostuu heikkojen sidosten, kuten vetysidosten ja ionisidosten kautta. Yhtym� muodostuu sit� helpommin mit� enemm�n n�it� sidoksia on; toisaalta sidoksia on sit� enemm�n mit� paremmin entsyymin aktiivinen kohta ja substraatti sopivat toisiinsa. Entsyymin valikoivuus selittyy t�t� taustaa vasten. Yksi substraatti sopii entsyymiin parhaiten, muut huonommin."
(Jyrki Heino, Matti Vuento, "Solubiologia", sivu 61) My�sk��n sidoksen lujuutta Spetnerin mallissa ei k�sitell�.
Spetnerin kaavan mukaan informaatiom��r� kasvaa sit� mukaa, mit� enemm�n on olemassa substraatteja, joihin entsyymi ei vaikuta, joten uusia substraatteja synnytt�m�ll� informaatiom��r� voisi kasvaa: T�m� johtaa my�s siihen ett� sin� hetken� kun entsyymi menett�� kokonaan toimintansa, sill� ei ole substraattia ja entsyymin informaatio nousee "Spetnerin tasolla valtavaksi" koska substraattien m��r� joihin entsyymi ei vaikuta on "kaikki mahdolliset". T�lt� kannalta eli�, jonka sis�ll� ei olisi ainuttakaan toimivaa entsyymi� sis�lt�isi "maksimaalisesti informaatiota". -Joskin juuri t�m� ominaisuus selitt�� miten "nukkuva geeni" voi informaatiota kadottamalla muuttua toiminnalliseksi. Ennusteena Spetnerin teoriassa on, ett� satunnaiset mutaatiot tuhoaisivat informaatiota, jolloin satunnaismutaatiory�ppy aina kasvattaisi "entsyymien laaja -alaisuutta" - jolloin koetta jatkettaessa lopulta jokainen entsyymi toimisi jokaisen substraatin kanssa.
Spetnerin tunnetuimmat s��nn�t ovat "satunnaiset muutokset eiv�t tuota informaatiota" ja ett� "mutaatiot karsivat informaatiota"
[] => v [] a // satunnaisia muutoksia, m��r�isyys ei kasva.
a valikoituu // v + [] katoavat, informaatio v�henee
a => a ai ab // satunnaisia muutoksia, m��r�isyys ei kasva
ab => valikoituu // informaatio v�henee
...
a...y => a...z, a..ya //satunnaisia muutoksia, m��r�isyys ei kasva
a...z => valikoituu // informaatio v�henee
Spetnerlogiikalla informaation m��r� on koko ajan v�hentynyt, vaikka se on kasvanut nollasta 123:n bittiin ja informaatio on eksaktia. Salakuljetettua informaatiota siin� on samassa suhteessa miss� luonnossakin. Algoritmi tiet�� ainoastaan kuinka hyv� satunnaisten mutaatioiden muuntelema ratkaisu on tiettyjen kriteereiden valossa(mahdollisimman pitk� aakkosj�rjestys). Samoin luonnossa ymp�r�iv�t luonnonlait m��r��v�t sen mitk� muodot s�ilyv�t ja mitk� eiv�t.
Evoluution potentiaali
"A classic example is to remove the gene that allows E. coli bacteria to digest lactose, but then give the colony of bacteria only lactose to eat. Of the millions in a colony, some bacteria re-evolve the ability to hydrolyze lactose and survive."
(Science Daily Barry G. Hallin tutkimuksesta, "Darwin's Time Machine: Scientists Begin Predicting Evolution's Next Step", 20.Maal.2002.)
Informaationtuotto -ongelmien varalle Spetner on kehitt�nyt "mikroevoluution potentiaaliin" eli "esiohjelmoitu kehitys". T�ss� "eli�t ovat hienos��detyt", jolloin ilmestyy "yll�tt�v��kin muuntelua".
H�n vetoaa usein Barry G. Hallin 1982 tekem��n tutkimukseen. Siin� osoitettiin, ett� populaatioon ilmestyy s��nn�nmukaisesti ja sangen nopeasti tietyt muutokset tietyss� valintapaineessa. Spetnerin tulkinta kyseisest� tutkimuksesta on, ett� hy�dylliset mutaatiot ovat liian yleisi�, jolloin ne eiv�t ole "aidosti informaatiota luovia"(~Spetnerin "truly beneficial") vaan kaivavat informaation "potentiaalistaan".
Barry Hall on suorittanut sittemmin t�ydent�vi� tutkimuksia, mutta n�ist� Spetner ei puhu. Onhan se toisaalta ik�v��, ett� jo "klassisessa esimerkiss�" ensin suoritetaan "deleetio", eli koko ominaisuuden poisto ja se silti kehittyy takaisin - suorastaan ennustettavan pilkuntarkasti. "Potentiaali" ei siis voi olla perim�ss�. Vaikuttaa "evoluution hyvyysfunktioilta" eli tutummin "luonnonvalinnalta": T�ll�in Spetnerin "mikroevoluution potentiaali" tarkoittaa yksinkertaisesti samaa asiaa, mihin viittaa my�s Ernst Meyer 2003 julkaistun kirjansa "Evoluutio" sivulla 157 lauseessa "Koska kaikki kuviteltavissa olevat hy�dylliset mutaatiot ovat jo valikoituneet vakaissa elinymp�rist�iss�, uusien hy�dyllisten mutaatioiden syntyminen on melko harvinaista." Eli evoluutiolla on olemassa (lokaali)optimi, jonka "yli tai ohi" informaatiota ei voi synty�. K�yt�nn�ss� Spetnerin "neodarwinismin kritiikki" lakkaa olemasta "neodarwinismin kritiikki�", jos h�n my�nt�� luonnonvalinnan t�m�n ominaisuuden, joten ainut vaihtoehto on vetoaminen "nukkuviin geeneihin". Eli pelkk� klassinen "sokerijuurikkaan sokeripitoisuusesimerkki" tai muut vastaavat, jossa osoitetaan, ett� "informaatiolla on lokaali optimi" ei riit�: Ollakseen "evoluutiolle ongelma" t�ytyy my�s pysty� erottamaan "lokaali optimi" ja "mikroevoluution koko kyky" toisistaan ja osoittamaan, ett� kyse ei ole pelk�st� "lokaalista optimista": Lokaali optimi -v�ite kun on ongelmallinen siksi ett� vaikka evoluutioalgoritmeissa voi lokaali optimi olla pysyv�, luonnossa "hyvyysfunktiot" muuttuvat ajan kanssa. Jopa hyvin yksinkertainen esimerkki osoittaa sen ett� hyvyysfunktiot ja "lokaalit optimit" evoluutiossa ovat pakotettuja muuttumaan, ne eiv�t ole mik��n sein� kuin korkeintaan v�liaikaisesti:
Oletetaan nyt ett� meill� on hyvyysfunktio jossa on lokaali optimi, joka ei kuitenkaan ole "korkein mahdollinen huippu". Sitten l�yd�mme organismin, joka kiipe�� siihen. Kun evoluutio saavuttaa maksimin, evoluutio lakkaa etenem�st�, jonka seurauksena juuri sen lokaalin optimin kohdalla alkaa olemaan ruuhkaa: Yksil�t kuluttavat samaa ravintoa, joka taas vaikuttaa hyvyysfunktioon. Seurauksena lokaalin optimin paikka muuttuu.
Lis�ksi evoluutio on moniulotteinen, joten pit�isi todistaa ett� optimi olisi rajattu joka suuntaan. Jos meill� on hyvyysavaruus, jolla on X ja Y akseli, ei voida sanoa ett� se ett� kohde on sellaisella kohdalla ett� sen Y on lokaalissa optimissa, se ei tarkoita ett� se ei voisi muuttua paremmaksi, se voi liikkua X:n kautta, ja t�m� X:n muutos voi jopa tarjota uuden kohdan, jossa Y ei en�� olekaan sidottu. (Eli lukkiutumia voi synty� ja ne voivat vaihtaa paikkaa.) "Lokaali optimi" ei siksi toimi "mikroevoluutioesteen� jota ei voi ylitt��".
Eli "hyvyysfunktiot" ovat tuskin koskaan staattisia ja "useampi ulottuvuus" tuo lis�ksi uusia kiertoteit�.
Toisin sanoen, Spetnerist� s��nn�nmukainen kehitys ei suinkaan osoita, ett� h�nen omat laskelmansa "rappeutumisen pakosta" ja "evoluution satunnaisuudesta eli kyvytt�myydest� tuottaa ennusteita", olisivat v��r�ss�. H�n ei my�sk��n p���ttele, ett� "informaation trendi on nouseva", vaan sen sijaan se kertoo siit�, ett� muuntelumahdollisuuksien m��r� on rajallinen ja n�ist� vaihtoehdoista valikoituu tietty "sattumanvaraisesti eli ymp�rist�st� riippuen". Samanaikaisesti, kun "aktuaalinen, havaittava informaatio" kasvaa, niin "potentiaali pienenee".
T�lt� kohdin on eritt�in vaikeaa n�hd� eroa evoluutikkojen n�kemykseen ; heist� luonnonvalinta valikoi hyv�t ja huonot ominaisuudet ja ett� "jossain on potentiaalisessa muodossa tieto siit� millaista informaatiota eli�iss� voi olla". -Barry Hallin lis�ksi mm. Richard Dawkins on kirjoittanut luonnonvalinnan tuottamasta ennustettavuudesta- Evoluutikoista universumin sis�lt�m�t luonnonlait sek� n�iden keskin�inen vaikutus ymp�rist�ss� sis�lt�v�t "potentiaalisessa muodossa" tiedon "mink�lainen eli� on siihen sopiva ja mik� v�hemm�n sopiva". Spetner tulkitsee osan informaatiosta "nukkuviksi geeneiksi", mutta ei rajaa t�t� ainoaksi vaihtoehdoksi. Sill� miss� "potentiaali -informaation" varasto on, ei ole merkitt�v� asia. "Mikroevoluution potentiaali" voidaan siis varsin hyvin ymm�rt�� my�s "teistiseksi evoluutioksi" eik� oikein mitenk��n "evoluutiokritiikiksi".
Toisin sanoen Spetner ei vetoa ulkoisiin rakenteisiin. H�nen n�kemyksens� kannalta on t�ysin mahdollista, ett� koko eli�kunta olisi l�ht�isin esimerkiksi pienen bakteerin "potentiaalista". T�m� tarkoittaa sit�, ett� Spetner voi ajatella nykyisest� fossiilirekordista sen, ett� "potentiaali -informaatio" selv�sti ajan kanssa muuttuu "aktuaaliseksi informaatioksi".
T�ll� n�kemyksell� on muutamia heikkouksia:
*Ensinn�kin mink�lainen tahansa fossiiliaineisto sopii Spetnerin n�kemykseen, eli fossiileilla n�kemyst� ei voi falsifioida.
*Toisaalta t�m� johtaa mu�s siihen, ett� "rappeutuneiden rakenteiden" esittely ei my�sk��n ole mik��n todiste : Informaatiom��r� kun ei ole pelk�st��n n�kyviss� rakenteissa.
*Kolmanneksi jos potentiaali olisi koodattuna perim��n, ja se vain "nousisi sielt� ajan mittaan esiin", pit�isi mallin selitt�� miten; Ongelmana kun on se, ett� eitoiminnallinen, piilossa oleva, potentiaali on kuitenkin altis satunnaisille mutaatioille, jonka pit�isi johtaa siihen ett� ilman valintapainetta potentiaali tuhoutuu pitk�ll� t�ht�imell�: Toisin sanoen siin� miss� evoluutio EI v�it� ett� ominaisuudet nousisivat esiin sattumalta, vaan p�invastoin korostavat luonnonvalinnan merkityst� siin� ett� toiminta ei tuhoudu, koodattuun, eitoiminnalliseen, potentiaaliin uskova joutuisi hyv�ksym��n "sattumalta esiin nousevan informaation". Potentiaali voisi toki nousta esiin "eisatunnaisin mutaatioin", mutta t�ll�isest� manipulaatiosta ei ole havaintoja.
Spetnerin malli koskee pelk�st��n entsyymeit�, eik� siksi ole sovellettavissa rakenteisiin tai mihink��n muuhun. Spetnerin mallin falsifiointi on kyseenalaista, koska periaatteessa mik� tahansa informaatio voi olla per�isin "jostakin tuntemattomasta potentiaalista" ja vaikka jokin mutaatio muuttaisi jotain entsyymi� "spesifisemm�ksi", on se ainoastaan "spesifisemm�ksi suhteessa tiet�miimme substraatteihin", jolloin ei voida tiet�� onko kyse "aidosta spesifioitumisesta" eli "spesifisyydest� suhteessa kaikkiin substraatteihin". Jos potentiaali katsotaan rajoitetuksi pelk�st��n perim��n, niin tilanne muuttuu. Spetner muuttuu empiriaksi heti, kun tapa mitata potentiaalin suuruus keksit��n, ja heti, kun Spetner onnistuu keksim��n sen mekanismin, joka takaa sen, ett� mutaatiot ovatkin hy�dyllisi�, informaatiota "potentiaalista" esiinkaivavia, sen sijaan ett� olisivat "satunnaisia".
Mutaatiot - uuden rakentaminen vai varastostahaku
"I agree with him that these mutations add information to the B-cell genome."
(Lee Spetner, "Lee Spetner/Edward Max Dialogue", 2006)
On havaittu sellaisia "satunnaisia" mutaatioita, jotka lis��v�t geneettist� informaatiota jopa Spetnerin menetelm�ll� mitattuna. Erityisen vaikeaksi Spetnerin mallille on ollut immuunisysteemi, koska vasta -aineet eiv�t ole yhdistettyn� "nukkuviin geeneihin", vaan ensimm�isell� kerralla kun tauti iskee, siihen sopivaa informaatiota ei ole, vaan se on generoitava satunnaisin mutaatioin. Spetner on my�nt�nyt sen, ett� immuunisysteemi lis�� informaatiota B -solun genomiin. Kyse ei siis ole "potentiaali - informaation esiinkaivamisesta", vaan aidosta informaation kasvusta. Eli jotain, jonka piti h�nen omista m��ritelmist��n k�sin olla mahdotonta.
Spetner esitt�� seuraavia syit� sille,miksi t�m� on mahdollista:
*Mutaatiom��r� on suuri : Arviot v�lill� 100 miljoonaa - 10 miljardia kappaletta.
T�m�n p��telm�n Spetner perustelee, sivulla 103, sill� ett� todenn�k�isyys tuottaa informaatiota "evoluutiosattumalta" saadaan, kun kerrotaan jokaisen v�livaiheen todenn�k�isyydet kesken��n : T�m� todenn�k�isyys taas saavutetaan arvaamalla tarpeeksi monta kertaa. T�ss� n�kemyksess� kummallista on:
-Luonnonvalintaa ei oteta huomioon lainkaan, jolloin v�livaiheiden k�ytt� itse asiassa menett�� merkityksens�. V�livaiheella voi olla merkityst� vain luonnonvalinnan kautta. "V�livaiheilla" luodaan ainoastaan valheellinen mielikuva "evoluution huomioon ottamisessa".
-Miten t�t� selityst� soveltaen informaatiota ei synny luonnossa "miljoonissa vuosissa", kun todenn�k�isyydet pysyv�t ja mutaatiot tuottavat jatkuvasti "arvauksia"? Eik� sen pit�isi olla pikemminkin v�ist�m�t�nt�?
On joka tapauksessa eritt�in erikoista, ett� Spetner, jotka kiist�� "aidosti hy�dyllisten mutaatioiden" olemassaolonkin kaikesta huolimatta itse tarjoaa mutaatioita "tarpeeksi isoissa m��riss� esiintyess��n" selityksen� "aidosti uuden informaation" synnylle. Eih�n "puhtaan todenn�k�isyyden" t�yttymiselle, sellaisenahan Spetner tilanteen laskee- ole ratkaisevaa se, kuinka lyhyess� ajassa yritykset tapahtuvat, vaan siin� kuinka monta kertaa n�in tapahtuu.
*Immuunisysteemiss� k�sitelt�v� informaatiom��r� on pieni
T�m� on "virhekatastrofirajaan" sovellettuna aivan j�rkev� argumentti. Suuret mutaatiom��r�t kuitenkin toimisivat hajottavana voimana. Perim�ss�mme mutaatiom��r� on selv�sti immuunisysteemi� pienempi informaatiom��r� ovat selv�sti suurempi. N�in suurempi informaatiom��r� joudutaan kompensoimaan, eik� virhekatastrofiin ajauduta.
*Systeemi on hienos��detty.
On aivan totta, ett� informaatio "on biteiss�": sek� informaation kasvu ett� sen v�heneminen ovat muutoksia niiss� - luokiteltiin informaatio mill� tavoin tahansa. N�in ollen pelk�t muutokset eiv�t riit�, vaan tarvitaan jotain, joka "tarkastaa" mitk� ovat hyvi� ja mitk� huonoja muutoksia. Immuunisysteemiss� on tapoja tunnistaa hyv�t ja huonot muutokset, Spetner kutstuu t�t� "hienos��d�ksi". Luonnossa vastaavaa ei kutsuta "hienos��d�ksi" vaan "luonnonvalinnaksi" - ja t�t� evoluutikot pit�v�t kovasti arvossa.
*Muutokset eiv�t tallennu perim��n.
Spetnerin k�sitys muuttuu t�ll� perusteella hieman kummalliseksi, koska h�nen v�itteens� on ymm�rrett�viss� "Informaatiota ei voi synty�, jos lopputulos tallennetaan." Lis�ongelmia t�lle tuo se, ett� vaikka onkin totta, ett� immuunisysteemi ei tallenna muutoksia ituradan perim��n, niin silti tiedot tallentuvat immuunisysteemin soluihin: syntyy resistenssi ja sama tauti paranee seuraavalla kerralla nopeammin. Ilmeisesti pelkk� tallennuskaan ei siis voi olla este, vaan ilmeisesti jokin vaikutus on sill� minne se tallennus tehd��n.. Tai sitten t�m� argumentti on n�enn�inen.
Kun n�m� kaikki kohdat otetaan huomioon on helpohko huomata, ett� immuunisysteemi on "analoginen" evoluutioprosesseihin. Evoluutiokin on hienostunut systeemi, jossa ymp�rist� asettaa rajaukset informaatiolle, ja jossa esiintyy runsaasti mutaatioita ja jossa on valintamekanismi.
Yhteenvetoa
"The NREH, on the other hand, postulates nonrandom variation. It does not fulfill the neo-Darwinian agenda in that it does not contribute to a natural explanation of the origin of life."
(Lee Spetner, "Not by Chance!", 1998, sivu 210)
Spetnerin keskittyy evoluutiokritiikkiin, mutta muista poiketen esitt�� my�s vastahypoteesin, "NREH":n.
*Evoluutiohypoteesissaan Spetner nojaa omaan informaatiom��rittelyyns�, joka ei ole kovin osuva -Spetner ei esimerkiksi k�sit�, ett� entsyymien ja substraattien sitoutumisella ei ole "kaikki tai ei mit��n" -luonnetta. H�n my�s laittaa yht�l�isyysmerkit "puhtaan Sattuman" ja "evoluution" v�liin. Spetnerin k�ytt�m� "hienos��t�perustelu", joka sallii informaation kasvun taas on ominaisuuksiltaan kovasti samanlainen, kuin luonnonvalinta.
*NREH:ssa keskeisint� on "evoluution ennalta ohjelmoitu potentiaali", joka m��r�� miten kehitys menee ja mink�laista informaatiota tulee. Spetner ei kuitenkaan selit� miten mutaatiot kaivavat potentiaalista informaatiota, eik� sit� miten t�t� potentiaalia voitaisiin tutkia. H�n paljastaa, ett� osa on "nukkuvia geenej�", mutta t�m� ei ole ainut menetelm�. Spetner ei my�sk��n kerro, syntyyk� "NREH":n piiriss� uutta informaatiota vai ei. Emme loppupeleiss� tied� mik� t�m� "potentiaali" on, emmek� my�sk��n mitk� ovat "NREH":n rajat, ja t�t� kautta kautta meill� ei ole keinoa falsifioida Spetnerin teoriaa: Esimerkiksi havainnot, joissa Spetnerin m��ritelm�n mukainen informaatio lis��ntyy ei falsifioi h�nen malliaan, koska ne voidaan tulkita "tuntemattomasta potentiaalista" johtuviksi. Spetner ei my�sk��n sovella evoluutioon soveltamaansa "Voisiko-se-toimia?" -kaavaansa omaan informaatiomalliinsa: l�hinn� koska h�nen mallinsa ei ainakaan toistaiseksi kerro mit��n sellaista mit� evoluutiokaan ei kertoisi - h�nen mallinsa keskeisimm�t, eli "mutaatioita ohjaava voima" ja "mikroevoluution potentiaali" kun ovat molemmat paitsi tuntemattomia my�s tuntemattoman suuruisia.
Kaikesta huolimatta Spetnerin malli ansaitsee erikoishuomion siit�, ett� se on toistaiseksi ainut kreationistien informaatiomalli, joka on sek� formalisaatio ett� vaihtoehtoinen malli.
Lis�tietoja:
Timo Tiainen,"Kriittinen arvio Spetnerin n�kemyksist�",2004
Ian Musgrave,"Information Theory and Creationism - Spetner and Biological Information",2005
Gert Korthof,"Could it work?",2003
Mark Chu-Carroll, "Using Bad Math to Create Bad Models to Produce Bad Results",2007
Spetner ei kuitenkaan koko h�nen kirjassaan k�sittele "Evoluution suuntaajaa" eli luonnonvalintaa muuten kuin muodossa "se karsii informaatiota", ottamatta huomioon "mink�laista informaatiota". N�inollen esimerkiksi Dawkinsin esitt�m�t "luonnonvalinnan rajoittamat, ennustettavat evoluutiopolut" j��v�t k�sittelem�tt� ja evoluutio k�sitell��n pelk�st��n "perinteisist� perinteisimm�ll�" evoluutio -olkinukkena, "puhtaansa sattumana". Parhaimmillaankin Spetner siis todistaa, ett� eli�kunta ei ole syntynyt kerralla sattumalta, mik� onkin aivan j�rkev��. Spetnerin n�kemys painottaa sit�, ett� mutaatiot ovat "eisatunnaisia", ja siksi hy�dyllisi� ja ett� "satunnaisuus ei informaatiota voisi tuottaa". Spetnerin suurin varsinainen ero "vallitsevaan uusdarwinismiin" onkin juuri se, ett� h�nen mallissaan valinta vaihtoehdoista on "a priori"(�lykk��n tahon koodaama "mikroevoluution potentiaali") ja evoluutiossa "a posteriori"(luonnonvalinta).
Todistaakseen n�kemyksens� oikeaksi, tulisi Spetnerin siis osoittaa miksi se kun joku �lykk��n tahon ohjelmoimat s��nn�t valitsevat "vapausasteisista systeemeist� funktionaalisesti sopivat", lis�� informaatiota, mutta jos luonnonvalinta tekee saman, niin informaation m��r�n kasvua ei voi tapahtua. T�m� voisi tapahtua esimerkiksi esitt�m�ll� tavan, jolla "potentiaalinen informaatiom��r�" voitaisiin selv�sti esiinsaada.
Spetnerin Informaatiom��rittely
"all point mutations that have been studied on the molecular level turn out to reduce the amount of information"
(Lee Spetner, "Not By Chance!", 1998, sivu 138)
Spetner soveltaa informaatiomalliaan entsyymeihin ja n�iden suhteeseen olemassaoleviin substraatteihin ja n�iden lukum��r��n. Spetnerin informaatiolla on kaksi piirrett�:
1: "odotusmittaus" ("expectation" measure) : Erilaisten merkkijonojen kokonaisuudella on v�hemm�n informaatiota kuin identtisten merkkijonojen kokonaisuudella.
2: "osoitemittaus" ("addressing" measure of information") : Pitemm�ss� merkkijonossa on enemm�n informaatiota. Informaatio on "kyll� tai ei"~"kaikki tai ei mit��n" -tyyppist�.
Spetnerin informaation m��r� kasvaa sit� suuremmaksi mit� enemm�n on substraatteja joihin entsyymi ei vaikuta. Eniten informaatiota on "spesifisess� entsyymiss�", joka tehoaa ainoastaan yhteen substraattiin.
M��rittely ja mitattavuus
"When I say a variation is not random, I mean that the chance of it occurring has something to do with how the organism adapts to its environment"
(Lee Spetner, "Not By Chance!", 1998, sivu 175)
Spetnerin m��rittelyn pohjalta luonnossa ilmenev�n informaation mittaaminen on mahdotonta, koska se vaatisi sen, ett� tutkittavan entsyymin toiminta tulisi testata suhteessa joka ikiseen universumissa mahdolliseen substraattiin : Ja n�iden lukum��r�� saati rakennetta emme edes tied�, saati sitten saa yhteiseen tilaan entsyymin kanssa.
Spetnerin informaatiomallissa oletetaan,ett� geeni toimii "joko-tai" mekanismilla ilman v�livaiheita; t�m�n ns. geeniperheet osoittavat v��r�ksi : niiss� useat samankaltaiset geenit hoitavat erilaisia, joskus osittain limitt�isi�kin, toimintoja. (Lokki, Saura & Tigerstedt,"Evoluutio ja populaatiot.",1986) N�inollen toiminto voi muuttua ja kun kahdentuminen on mahdollista, voi kahdentuma + toisen muutos johtaa uuden informaation syntyyn "eritt�in mikroevolutiivisesti".
Spetnerin informaatiom��rittelyss� on my�skin joitain puutteita. Siin� ei esimerkiksi oteta lainkaan huomioon toimintalaatua. Jos entsyymi toimii mitenk��n on se informaatiota; Spetnerin informaatio on "joko tai", vaikka entsyymien toiminta ei t�ll�ist� ole, ja eli�n toiminnallisuuden kannalta on kuitenkin suunnatonta merkityst� sill� mill� teholla entsyymi toimii."Entsyymi - substraattiyhdistym� muodostuu heikkojen sidosten, kuten vetysidosten ja ionisidosten kautta. Yhtym� muodostuu sit� helpommin mit� enemm�n n�it� sidoksia on; toisaalta sidoksia on sit� enemm�n mit� paremmin entsyymin aktiivinen kohta ja substraatti sopivat toisiinsa. Entsyymin valikoivuus selittyy t�t� taustaa vasten. Yksi substraatti sopii entsyymiin parhaiten, muut huonommin."
(Jyrki Heino, Matti Vuento, "Solubiologia", sivu 61) My�sk��n sidoksen lujuutta Spetnerin mallissa ei k�sitell�.
Spetnerin kaavan mukaan informaatiom��r� kasvaa sit� mukaa, mit� enemm�n on olemassa substraatteja, joihin entsyymi ei vaikuta, joten uusia substraatteja synnytt�m�ll� informaatiom��r� voisi kasvaa: T�m� johtaa my�s siihen ett� sin� hetken� kun entsyymi menett�� kokonaan toimintansa, sill� ei ole substraattia ja entsyymin informaatio nousee "Spetnerin tasolla valtavaksi" koska substraattien m��r� joihin entsyymi ei vaikuta on "kaikki mahdolliset". T�lt� kannalta eli�, jonka sis�ll� ei olisi ainuttakaan toimivaa entsyymi� sis�lt�isi "maksimaalisesti informaatiota". -Joskin juuri t�m� ominaisuus selitt�� miten "nukkuva geeni" voi informaatiota kadottamalla muuttua toiminnalliseksi. Ennusteena Spetnerin teoriassa on, ett� satunnaiset mutaatiot tuhoaisivat informaatiota, jolloin satunnaismutaatiory�ppy aina kasvattaisi "entsyymien laaja -alaisuutta" - jolloin koetta jatkettaessa lopulta jokainen entsyymi toimisi jokaisen substraatin kanssa.
Spetnerin tunnetuimmat s��nn�t ovat "satunnaiset muutokset eiv�t tuota informaatiota" ja ett� "mutaatiot karsivat informaatiota"
[] => v [] a // satunnaisia muutoksia, m��r�isyys ei kasva.
a valikoituu // v + [] katoavat, informaatio v�henee
a => a ai ab // satunnaisia muutoksia, m��r�isyys ei kasva
ab => valikoituu // informaatio v�henee
...
a...y => a...z, a..ya //satunnaisia muutoksia, m��r�isyys ei kasva
a...z => valikoituu // informaatio v�henee
Spetnerlogiikalla informaation m��r� on koko ajan v�hentynyt, vaikka se on kasvanut nollasta 123:n bittiin ja informaatio on eksaktia. Salakuljetettua informaatiota siin� on samassa suhteessa miss� luonnossakin. Algoritmi tiet�� ainoastaan kuinka hyv� satunnaisten mutaatioiden muuntelema ratkaisu on tiettyjen kriteereiden valossa(mahdollisimman pitk� aakkosj�rjestys). Samoin luonnossa ymp�r�iv�t luonnonlait m��r��v�t sen mitk� muodot s�ilyv�t ja mitk� eiv�t.
Evoluution potentiaali
"A classic example is to remove the gene that allows E. coli bacteria to digest lactose, but then give the colony of bacteria only lactose to eat. Of the millions in a colony, some bacteria re-evolve the ability to hydrolyze lactose and survive."
(Science Daily Barry G. Hallin tutkimuksesta, "Darwin's Time Machine: Scientists Begin Predicting Evolution's Next Step", 20.Maal.2002.)
Informaationtuotto -ongelmien varalle Spetner on kehitt�nyt "mikroevoluution potentiaaliin" eli "esiohjelmoitu kehitys". T�ss� "eli�t ovat hienos��detyt", jolloin ilmestyy "yll�tt�v��kin muuntelua".
H�n vetoaa usein Barry G. Hallin 1982 tekem��n tutkimukseen. Siin� osoitettiin, ett� populaatioon ilmestyy s��nn�nmukaisesti ja sangen nopeasti tietyt muutokset tietyss� valintapaineessa. Spetnerin tulkinta kyseisest� tutkimuksesta on, ett� hy�dylliset mutaatiot ovat liian yleisi�, jolloin ne eiv�t ole "aidosti informaatiota luovia"(~Spetnerin "truly beneficial") vaan kaivavat informaation "potentiaalistaan".
Barry Hall on suorittanut sittemmin t�ydent�vi� tutkimuksia, mutta n�ist� Spetner ei puhu. Onhan se toisaalta ik�v��, ett� jo "klassisessa esimerkiss�" ensin suoritetaan "deleetio", eli koko ominaisuuden poisto ja se silti kehittyy takaisin - suorastaan ennustettavan pilkuntarkasti. "Potentiaali" ei siis voi olla perim�ss�. Vaikuttaa "evoluution hyvyysfunktioilta" eli tutummin "luonnonvalinnalta": T�ll�in Spetnerin "mikroevoluution potentiaali" tarkoittaa yksinkertaisesti samaa asiaa, mihin viittaa my�s Ernst Meyer 2003 julkaistun kirjansa "Evoluutio" sivulla 157 lauseessa "Koska kaikki kuviteltavissa olevat hy�dylliset mutaatiot ovat jo valikoituneet vakaissa elinymp�rist�iss�, uusien hy�dyllisten mutaatioiden syntyminen on melko harvinaista." Eli evoluutiolla on olemassa (lokaali)optimi, jonka "yli tai ohi" informaatiota ei voi synty�. K�yt�nn�ss� Spetnerin "neodarwinismin kritiikki" lakkaa olemasta "neodarwinismin kritiikki�", jos h�n my�nt�� luonnonvalinnan t�m�n ominaisuuden, joten ainut vaihtoehto on vetoaminen "nukkuviin geeneihin". Eli pelkk� klassinen "sokerijuurikkaan sokeripitoisuusesimerkki" tai muut vastaavat, jossa osoitetaan, ett� "informaatiolla on lokaali optimi" ei riit�: Ollakseen "evoluutiolle ongelma" t�ytyy my�s pysty� erottamaan "lokaali optimi" ja "mikroevoluution koko kyky" toisistaan ja osoittamaan, ett� kyse ei ole pelk�st� "lokaalista optimista": Lokaali optimi -v�ite kun on ongelmallinen siksi ett� vaikka evoluutioalgoritmeissa voi lokaali optimi olla pysyv�, luonnossa "hyvyysfunktiot" muuttuvat ajan kanssa. Jopa hyvin yksinkertainen esimerkki osoittaa sen ett� hyvyysfunktiot ja "lokaalit optimit" evoluutiossa ovat pakotettuja muuttumaan, ne eiv�t ole mik��n sein� kuin korkeintaan v�liaikaisesti:
Oletetaan nyt ett� meill� on hyvyysfunktio jossa on lokaali optimi, joka ei kuitenkaan ole "korkein mahdollinen huippu". Sitten l�yd�mme organismin, joka kiipe�� siihen. Kun evoluutio saavuttaa maksimin, evoluutio lakkaa etenem�st�, jonka seurauksena juuri sen lokaalin optimin kohdalla alkaa olemaan ruuhkaa: Yksil�t kuluttavat samaa ravintoa, joka taas vaikuttaa hyvyysfunktioon. Seurauksena lokaalin optimin paikka muuttuu.
Lis�ksi evoluutio on moniulotteinen, joten pit�isi todistaa ett� optimi olisi rajattu joka suuntaan. Jos meill� on hyvyysavaruus, jolla on X ja Y akseli, ei voida sanoa ett� se ett� kohde on sellaisella kohdalla ett� sen Y on lokaalissa optimissa, se ei tarkoita ett� se ei voisi muuttua paremmaksi, se voi liikkua X:n kautta, ja t�m� X:n muutos voi jopa tarjota uuden kohdan, jossa Y ei en�� olekaan sidottu. (Eli lukkiutumia voi synty� ja ne voivat vaihtaa paikkaa.) "Lokaali optimi" ei siksi toimi "mikroevoluutioesteen� jota ei voi ylitt��".
Eli "hyvyysfunktiot" ovat tuskin koskaan staattisia ja "useampi ulottuvuus" tuo lis�ksi uusia kiertoteit�.
Toisin sanoen, Spetnerist� s��nn�nmukainen kehitys ei suinkaan osoita, ett� h�nen omat laskelmansa "rappeutumisen pakosta" ja "evoluution satunnaisuudesta eli kyvytt�myydest� tuottaa ennusteita", olisivat v��r�ss�. H�n ei my�sk��n p���ttele, ett� "informaation trendi on nouseva", vaan sen sijaan se kertoo siit�, ett� muuntelumahdollisuuksien m��r� on rajallinen ja n�ist� vaihtoehdoista valikoituu tietty "sattumanvaraisesti eli ymp�rist�st� riippuen". Samanaikaisesti, kun "aktuaalinen, havaittava informaatio" kasvaa, niin "potentiaali pienenee".
T�lt� kohdin on eritt�in vaikeaa n�hd� eroa evoluutikkojen n�kemykseen ; heist� luonnonvalinta valikoi hyv�t ja huonot ominaisuudet ja ett� "jossain on potentiaalisessa muodossa tieto siit� millaista informaatiota eli�iss� voi olla". -Barry Hallin lis�ksi mm. Richard Dawkins on kirjoittanut luonnonvalinnan tuottamasta ennustettavuudesta- Evoluutikoista universumin sis�lt�m�t luonnonlait sek� n�iden keskin�inen vaikutus ymp�rist�ss� sis�lt�v�t "potentiaalisessa muodossa" tiedon "mink�lainen eli� on siihen sopiva ja mik� v�hemm�n sopiva". Spetner tulkitsee osan informaatiosta "nukkuviksi geeneiksi", mutta ei rajaa t�t� ainoaksi vaihtoehdoksi. Sill� miss� "potentiaali -informaation" varasto on, ei ole merkitt�v� asia. "Mikroevoluution potentiaali" voidaan siis varsin hyvin ymm�rt�� my�s "teistiseksi evoluutioksi" eik� oikein mitenk��n "evoluutiokritiikiksi".
Toisin sanoen Spetner ei vetoa ulkoisiin rakenteisiin. H�nen n�kemyksens� kannalta on t�ysin mahdollista, ett� koko eli�kunta olisi l�ht�isin esimerkiksi pienen bakteerin "potentiaalista". T�m� tarkoittaa sit�, ett� Spetner voi ajatella nykyisest� fossiilirekordista sen, ett� "potentiaali -informaatio" selv�sti ajan kanssa muuttuu "aktuaaliseksi informaatioksi".
T�ll� n�kemyksell� on muutamia heikkouksia:
*Ensinn�kin mink�lainen tahansa fossiiliaineisto sopii Spetnerin n�kemykseen, eli fossiileilla n�kemyst� ei voi falsifioida.
*Toisaalta t�m� johtaa mu�s siihen, ett� "rappeutuneiden rakenteiden" esittely ei my�sk��n ole mik��n todiste : Informaatiom��r� kun ei ole pelk�st��n n�kyviss� rakenteissa.
*Kolmanneksi jos potentiaali olisi koodattuna perim��n, ja se vain "nousisi sielt� ajan mittaan esiin", pit�isi mallin selitt�� miten; Ongelmana kun on se, ett� eitoiminnallinen, piilossa oleva, potentiaali on kuitenkin altis satunnaisille mutaatioille, jonka pit�isi johtaa siihen ett� ilman valintapainetta potentiaali tuhoutuu pitk�ll� t�ht�imell�: Toisin sanoen siin� miss� evoluutio EI v�it� ett� ominaisuudet nousisivat esiin sattumalta, vaan p�invastoin korostavat luonnonvalinnan merkityst� siin� ett� toiminta ei tuhoudu, koodattuun, eitoiminnalliseen, potentiaaliin uskova joutuisi hyv�ksym��n "sattumalta esiin nousevan informaation". Potentiaali voisi toki nousta esiin "eisatunnaisin mutaatioin", mutta t�ll�isest� manipulaatiosta ei ole havaintoja.
Spetnerin malli koskee pelk�st��n entsyymeit�, eik� siksi ole sovellettavissa rakenteisiin tai mihink��n muuhun. Spetnerin mallin falsifiointi on kyseenalaista, koska periaatteessa mik� tahansa informaatio voi olla per�isin "jostakin tuntemattomasta potentiaalista" ja vaikka jokin mutaatio muuttaisi jotain entsyymi� "spesifisemm�ksi", on se ainoastaan "spesifisemm�ksi suhteessa tiet�miimme substraatteihin", jolloin ei voida tiet�� onko kyse "aidosta spesifioitumisesta" eli "spesifisyydest� suhteessa kaikkiin substraatteihin". Jos potentiaali katsotaan rajoitetuksi pelk�st��n perim��n, niin tilanne muuttuu. Spetner muuttuu empiriaksi heti, kun tapa mitata potentiaalin suuruus keksit��n, ja heti, kun Spetner onnistuu keksim��n sen mekanismin, joka takaa sen, ett� mutaatiot ovatkin hy�dyllisi�, informaatiota "potentiaalista" esiinkaivavia, sen sijaan ett� olisivat "satunnaisia".
Mutaatiot - uuden rakentaminen vai varastostahaku
"I agree with him that these mutations add information to the B-cell genome."
(Lee Spetner, "Lee Spetner/Edward Max Dialogue", 2006)
On havaittu sellaisia "satunnaisia" mutaatioita, jotka lis��v�t geneettist� informaatiota jopa Spetnerin menetelm�ll� mitattuna. Erityisen vaikeaksi Spetnerin mallille on ollut immuunisysteemi, koska vasta -aineet eiv�t ole yhdistettyn� "nukkuviin geeneihin", vaan ensimm�isell� kerralla kun tauti iskee, siihen sopivaa informaatiota ei ole, vaan se on generoitava satunnaisin mutaatioin. Spetner on my�nt�nyt sen, ett� immuunisysteemi lis�� informaatiota B -solun genomiin. Kyse ei siis ole "potentiaali - informaation esiinkaivamisesta", vaan aidosta informaation kasvusta. Eli jotain, jonka piti h�nen omista m��ritelmist��n k�sin olla mahdotonta.
Spetner esitt�� seuraavia syit� sille,miksi t�m� on mahdollista:
*Mutaatiom��r� on suuri : Arviot v�lill� 100 miljoonaa - 10 miljardia kappaletta.
T�m�n p��telm�n Spetner perustelee, sivulla 103, sill� ett� todenn�k�isyys tuottaa informaatiota "evoluutiosattumalta" saadaan, kun kerrotaan jokaisen v�livaiheen todenn�k�isyydet kesken��n : T�m� todenn�k�isyys taas saavutetaan arvaamalla tarpeeksi monta kertaa. T�ss� n�kemyksess� kummallista on:
-Luonnonvalintaa ei oteta huomioon lainkaan, jolloin v�livaiheiden k�ytt� itse asiassa menett�� merkityksens�. V�livaiheella voi olla merkityst� vain luonnonvalinnan kautta. "V�livaiheilla" luodaan ainoastaan valheellinen mielikuva "evoluution huomioon ottamisessa".
-Miten t�t� selityst� soveltaen informaatiota ei synny luonnossa "miljoonissa vuosissa", kun todenn�k�isyydet pysyv�t ja mutaatiot tuottavat jatkuvasti "arvauksia"? Eik� sen pit�isi olla pikemminkin v�ist�m�t�nt�?
On joka tapauksessa eritt�in erikoista, ett� Spetner, jotka kiist�� "aidosti hy�dyllisten mutaatioiden" olemassaolonkin kaikesta huolimatta itse tarjoaa mutaatioita "tarpeeksi isoissa m��riss� esiintyess��n" selityksen� "aidosti uuden informaation" synnylle. Eih�n "puhtaan todenn�k�isyyden" t�yttymiselle, sellaisenahan Spetner tilanteen laskee- ole ratkaisevaa se, kuinka lyhyess� ajassa yritykset tapahtuvat, vaan siin� kuinka monta kertaa n�in tapahtuu.
*Immuunisysteemiss� k�sitelt�v� informaatiom��r� on pieni
T�m� on "virhekatastrofirajaan" sovellettuna aivan j�rkev� argumentti. Suuret mutaatiom��r�t kuitenkin toimisivat hajottavana voimana. Perim�ss�mme mutaatiom��r� on selv�sti immuunisysteemi� pienempi informaatiom��r� ovat selv�sti suurempi. N�in suurempi informaatiom��r� joudutaan kompensoimaan, eik� virhekatastrofiin ajauduta.
*Systeemi on hienos��detty.
On aivan totta, ett� informaatio "on biteiss�": sek� informaation kasvu ett� sen v�heneminen ovat muutoksia niiss� - luokiteltiin informaatio mill� tavoin tahansa. N�in ollen pelk�t muutokset eiv�t riit�, vaan tarvitaan jotain, joka "tarkastaa" mitk� ovat hyvi� ja mitk� huonoja muutoksia. Immuunisysteemiss� on tapoja tunnistaa hyv�t ja huonot muutokset, Spetner kutstuu t�t� "hienos��d�ksi". Luonnossa vastaavaa ei kutsuta "hienos��d�ksi" vaan "luonnonvalinnaksi" - ja t�t� evoluutikot pit�v�t kovasti arvossa.
*Muutokset eiv�t tallennu perim��n.
Spetnerin k�sitys muuttuu t�ll� perusteella hieman kummalliseksi, koska h�nen v�itteens� on ymm�rrett�viss� "Informaatiota ei voi synty�, jos lopputulos tallennetaan." Lis�ongelmia t�lle tuo se, ett� vaikka onkin totta, ett� immuunisysteemi ei tallenna muutoksia ituradan perim��n, niin silti tiedot tallentuvat immuunisysteemin soluihin: syntyy resistenssi ja sama tauti paranee seuraavalla kerralla nopeammin. Ilmeisesti pelkk� tallennuskaan ei siis voi olla este, vaan ilmeisesti jokin vaikutus on sill� minne se tallennus tehd��n.. Tai sitten t�m� argumentti on n�enn�inen.
Kun n�m� kaikki kohdat otetaan huomioon on helpohko huomata, ett� immuunisysteemi on "analoginen" evoluutioprosesseihin. Evoluutiokin on hienostunut systeemi, jossa ymp�rist� asettaa rajaukset informaatiolle, ja jossa esiintyy runsaasti mutaatioita ja jossa on valintamekanismi.
Yhteenvetoa
"The NREH, on the other hand, postulates nonrandom variation. It does not fulfill the neo-Darwinian agenda in that it does not contribute to a natural explanation of the origin of life."
(Lee Spetner, "Not by Chance!", 1998, sivu 210)
Spetnerin keskittyy evoluutiokritiikkiin, mutta muista poiketen esitt�� my�s vastahypoteesin, "NREH":n.
*Evoluutiohypoteesissaan Spetner nojaa omaan informaatiom��rittelyyns�, joka ei ole kovin osuva -Spetner ei esimerkiksi k�sit�, ett� entsyymien ja substraattien sitoutumisella ei ole "kaikki tai ei mit��n" -luonnetta. H�n my�s laittaa yht�l�isyysmerkit "puhtaan Sattuman" ja "evoluution" v�liin. Spetnerin k�ytt�m� "hienos��t�perustelu", joka sallii informaation kasvun taas on ominaisuuksiltaan kovasti samanlainen, kuin luonnonvalinta.
*NREH:ssa keskeisint� on "evoluution ennalta ohjelmoitu potentiaali", joka m��r�� miten kehitys menee ja mink�laista informaatiota tulee. Spetner ei kuitenkaan selit� miten mutaatiot kaivavat potentiaalista informaatiota, eik� sit� miten t�t� potentiaalia voitaisiin tutkia. H�n paljastaa, ett� osa on "nukkuvia geenej�", mutta t�m� ei ole ainut menetelm�. Spetner ei my�sk��n kerro, syntyyk� "NREH":n piiriss� uutta informaatiota vai ei. Emme loppupeleiss� tied� mik� t�m� "potentiaali" on, emmek� my�sk��n mitk� ovat "NREH":n rajat, ja t�t� kautta kautta meill� ei ole keinoa falsifioida Spetnerin teoriaa: Esimerkiksi havainnot, joissa Spetnerin m��ritelm�n mukainen informaatio lis��ntyy ei falsifioi h�nen malliaan, koska ne voidaan tulkita "tuntemattomasta potentiaalista" johtuviksi. Spetner ei my�sk��n sovella evoluutioon soveltamaansa "Voisiko-se-toimia?" -kaavaansa omaan informaatiomalliinsa: l�hinn� koska h�nen mallinsa ei ainakaan toistaiseksi kerro mit��n sellaista mit� evoluutiokaan ei kertoisi - h�nen mallinsa keskeisimm�t, eli "mutaatioita ohjaava voima" ja "mikroevoluution potentiaali" kun ovat molemmat paitsi tuntemattomia my�s tuntemattoman suuruisia.
Kaikesta huolimatta Spetnerin malli ansaitsee erikoishuomion siit�, ett� se on toistaiseksi ainut kreationistien informaatiomalli, joka on sek� formalisaatio ett� vaihtoehtoinen malli.
Lis�tietoja:
Timo Tiainen,"Kriittinen arvio Spetnerin n�kemyksist�",2004
Ian Musgrave,"Information Theory and Creationism - Spetner and Biological Information",2005
Gert Korthof,"Could it work?",2003
Mark Chu-Carroll, "Using Bad Math to Create Bad Models to Produce Bad Results",2007
(Lee Spetner, "Not By Chance!", 1998, sivu 138)
Spetner soveltaa informaatiomalliaan entsyymeihin ja n�iden suhteeseen olemassaoleviin substraatteihin ja n�iden lukum��r��n. Spetnerin informaatiolla on kaksi piirrett�:
1: "odotusmittaus" ("expectation" measure) : Erilaisten merkkijonojen kokonaisuudella on v�hemm�n informaatiota kuin identtisten merkkijonojen kokonaisuudella.
2: "osoitemittaus" ("addressing" measure of information") : Pitemm�ss� merkkijonossa on enemm�n informaatiota. Informaatio on "kyll� tai ei"~"kaikki tai ei mit��n" -tyyppist�.
Spetnerin informaation m��r� kasvaa sit� suuremmaksi mit� enemm�n on substraatteja joihin entsyymi ei vaikuta. Eniten informaatiota on "spesifisess� entsyymiss�", joka tehoaa ainoastaan yhteen substraattiin.
M��rittely ja mitattavuus
"When I say a variation is not random, I mean that the chance of it occurring has something to do with how the organism adapts to its environment"
(Lee Spetner, "Not By Chance!", 1998, sivu 175)
Spetnerin m��rittelyn pohjalta luonnossa ilmenev�n informaation mittaaminen on mahdotonta, koska se vaatisi sen, ett� tutkittavan entsyymin toiminta tulisi testata suhteessa joka ikiseen universumissa mahdolliseen substraattiin : Ja n�iden lukum��r�� saati rakennetta emme edes tied�, saati sitten saa yhteiseen tilaan entsyymin kanssa.
Spetnerin informaatiomallissa oletetaan,ett� geeni toimii "joko-tai" mekanismilla ilman v�livaiheita; t�m�n ns. geeniperheet osoittavat v��r�ksi : niiss� useat samankaltaiset geenit hoitavat erilaisia, joskus osittain limitt�isi�kin, toimintoja. (Lokki, Saura & Tigerstedt,"Evoluutio ja populaatiot.",1986) N�inollen toiminto voi muuttua ja kun kahdentuminen on mahdollista, voi kahdentuma + toisen muutos johtaa uuden informaation syntyyn "eritt�in mikroevolutiivisesti".
Spetnerin informaatiom��rittelyss� on my�skin joitain puutteita. Siin� ei esimerkiksi oteta lainkaan huomioon toimintalaatua. Jos entsyymi toimii mitenk��n on se informaatiota; Spetnerin informaatio on "joko tai", vaikka entsyymien toiminta ei t�ll�ist� ole, ja eli�n toiminnallisuuden kannalta on kuitenkin suunnatonta merkityst� sill� mill� teholla entsyymi toimii."Entsyymi - substraattiyhdistym� muodostuu heikkojen sidosten, kuten vetysidosten ja ionisidosten kautta. Yhtym� muodostuu sit� helpommin mit� enemm�n n�it� sidoksia on; toisaalta sidoksia on sit� enemm�n mit� paremmin entsyymin aktiivinen kohta ja substraatti sopivat toisiinsa. Entsyymin valikoivuus selittyy t�t� taustaa vasten. Yksi substraatti sopii entsyymiin parhaiten, muut huonommin."
(Jyrki Heino, Matti Vuento, "Solubiologia", sivu 61) My�sk��n sidoksen lujuutta Spetnerin mallissa ei k�sitell�.
Spetnerin kaavan mukaan informaatiom��r� kasvaa sit� mukaa, mit� enemm�n on olemassa substraatteja, joihin entsyymi ei vaikuta, joten uusia substraatteja synnytt�m�ll� informaatiom��r� voisi kasvaa: T�m� johtaa my�s siihen ett� sin� hetken� kun entsyymi menett�� kokonaan toimintansa, sill� ei ole substraattia ja entsyymin informaatio nousee "Spetnerin tasolla valtavaksi" koska substraattien m��r� joihin entsyymi ei vaikuta on "kaikki mahdolliset". T�lt� kannalta eli�, jonka sis�ll� ei olisi ainuttakaan toimivaa entsyymi� sis�lt�isi "maksimaalisesti informaatiota". -Joskin juuri t�m� ominaisuus selitt�� miten "nukkuva geeni" voi informaatiota kadottamalla muuttua toiminnalliseksi. Ennusteena Spetnerin teoriassa on, ett� satunnaiset mutaatiot tuhoaisivat informaatiota, jolloin satunnaismutaatiory�ppy aina kasvattaisi "entsyymien laaja -alaisuutta" - jolloin koetta jatkettaessa lopulta jokainen entsyymi toimisi jokaisen substraatin kanssa.
Spetnerin tunnetuimmat s��nn�t ovat "satunnaiset muutokset eiv�t tuota informaatiota" ja ett� "mutaatiot karsivat informaatiota"
[] => v [] a // satunnaisia muutoksia, m��r�isyys ei kasva.
a valikoituu // v + [] katoavat, informaatio v�henee
a => a ai ab // satunnaisia muutoksia, m��r�isyys ei kasva
ab => valikoituu // informaatio v�henee
...
a...y => a...z, a..ya //satunnaisia muutoksia, m��r�isyys ei kasva
a...z => valikoituu // informaatio v�henee
Spetnerlogiikalla informaation m��r� on koko ajan v�hentynyt, vaikka se on kasvanut nollasta 123:n bittiin ja informaatio on eksaktia. Salakuljetettua informaatiota siin� on samassa suhteessa miss� luonnossakin. Algoritmi tiet�� ainoastaan kuinka hyv� satunnaisten mutaatioiden muuntelema ratkaisu on tiettyjen kriteereiden valossa(mahdollisimman pitk� aakkosj�rjestys). Samoin luonnossa ymp�r�iv�t luonnonlait m��r��v�t sen mitk� muodot s�ilyv�t ja mitk� eiv�t.
Evoluution potentiaali
"A classic example is to remove the gene that allows E. coli bacteria to digest lactose, but then give the colony of bacteria only lactose to eat. Of the millions in a colony, some bacteria re-evolve the ability to hydrolyze lactose and survive."
(Science Daily Barry G. Hallin tutkimuksesta, "Darwin's Time Machine: Scientists Begin Predicting Evolution's Next Step", 20.Maal.2002.)
Informaationtuotto -ongelmien varalle Spetner on kehitt�nyt "mikroevoluution potentiaaliin" eli "esiohjelmoitu kehitys". T�ss� "eli�t ovat hienos��detyt", jolloin ilmestyy "yll�tt�v��kin muuntelua".
H�n vetoaa usein Barry G. Hallin 1982 tekem��n tutkimukseen. Siin� osoitettiin, ett� populaatioon ilmestyy s��nn�nmukaisesti ja sangen nopeasti tietyt muutokset tietyss� valintapaineessa. Spetnerin tulkinta kyseisest� tutkimuksesta on, ett� hy�dylliset mutaatiot ovat liian yleisi�, jolloin ne eiv�t ole "aidosti informaatiota luovia"(~Spetnerin "truly beneficial") vaan kaivavat informaation "potentiaalistaan".
Barry Hall on suorittanut sittemmin t�ydent�vi� tutkimuksia, mutta n�ist� Spetner ei puhu. Onhan se toisaalta ik�v��, ett� jo "klassisessa esimerkiss�" ensin suoritetaan "deleetio", eli koko ominaisuuden poisto ja se silti kehittyy takaisin - suorastaan ennustettavan pilkuntarkasti. "Potentiaali" ei siis voi olla perim�ss�. Vaikuttaa "evoluution hyvyysfunktioilta" eli tutummin "luonnonvalinnalta": T�ll�in Spetnerin "mikroevoluution potentiaali" tarkoittaa yksinkertaisesti samaa asiaa, mihin viittaa my�s Ernst Meyer 2003 julkaistun kirjansa "Evoluutio" sivulla 157 lauseessa "Koska kaikki kuviteltavissa olevat hy�dylliset mutaatiot ovat jo valikoituneet vakaissa elinymp�rist�iss�, uusien hy�dyllisten mutaatioiden syntyminen on melko harvinaista." Eli evoluutiolla on olemassa (lokaali)optimi, jonka "yli tai ohi" informaatiota ei voi synty�. K�yt�nn�ss� Spetnerin "neodarwinismin kritiikki" lakkaa olemasta "neodarwinismin kritiikki�", jos h�n my�nt�� luonnonvalinnan t�m�n ominaisuuden, joten ainut vaihtoehto on vetoaminen "nukkuviin geeneihin". Eli pelkk� klassinen "sokerijuurikkaan sokeripitoisuusesimerkki" tai muut vastaavat, jossa osoitetaan, ett� "informaatiolla on lokaali optimi" ei riit�: Ollakseen "evoluutiolle ongelma" t�ytyy my�s pysty� erottamaan "lokaali optimi" ja "mikroevoluution koko kyky" toisistaan ja osoittamaan, ett� kyse ei ole pelk�st� "lokaalista optimista": Lokaali optimi -v�ite kun on ongelmallinen siksi ett� vaikka evoluutioalgoritmeissa voi lokaali optimi olla pysyv�, luonnossa "hyvyysfunktiot" muuttuvat ajan kanssa. Jopa hyvin yksinkertainen esimerkki osoittaa sen ett� hyvyysfunktiot ja "lokaalit optimit" evoluutiossa ovat pakotettuja muuttumaan, ne eiv�t ole mik��n sein� kuin korkeintaan v�liaikaisesti:
Oletetaan nyt ett� meill� on hyvyysfunktio jossa on lokaali optimi, joka ei kuitenkaan ole "korkein mahdollinen huippu". Sitten l�yd�mme organismin, joka kiipe�� siihen. Kun evoluutio saavuttaa maksimin, evoluutio lakkaa etenem�st�, jonka seurauksena juuri sen lokaalin optimin kohdalla alkaa olemaan ruuhkaa: Yksil�t kuluttavat samaa ravintoa, joka taas vaikuttaa hyvyysfunktioon. Seurauksena lokaalin optimin paikka muuttuu.
Lis�ksi evoluutio on moniulotteinen, joten pit�isi todistaa ett� optimi olisi rajattu joka suuntaan. Jos meill� on hyvyysavaruus, jolla on X ja Y akseli, ei voida sanoa ett� se ett� kohde on sellaisella kohdalla ett� sen Y on lokaalissa optimissa, se ei tarkoita ett� se ei voisi muuttua paremmaksi, se voi liikkua X:n kautta, ja t�m� X:n muutos voi jopa tarjota uuden kohdan, jossa Y ei en�� olekaan sidottu. (Eli lukkiutumia voi synty� ja ne voivat vaihtaa paikkaa.) "Lokaali optimi" ei siksi toimi "mikroevoluutioesteen� jota ei voi ylitt��".
Eli "hyvyysfunktiot" ovat tuskin koskaan staattisia ja "useampi ulottuvuus" tuo lis�ksi uusia kiertoteit�.
Toisin sanoen, Spetnerist� s��nn�nmukainen kehitys ei suinkaan osoita, ett� h�nen omat laskelmansa "rappeutumisen pakosta" ja "evoluution satunnaisuudesta eli kyvytt�myydest� tuottaa ennusteita", olisivat v��r�ss�. H�n ei my�sk��n p���ttele, ett� "informaation trendi on nouseva", vaan sen sijaan se kertoo siit�, ett� muuntelumahdollisuuksien m��r� on rajallinen ja n�ist� vaihtoehdoista valikoituu tietty "sattumanvaraisesti eli ymp�rist�st� riippuen". Samanaikaisesti, kun "aktuaalinen, havaittava informaatio" kasvaa, niin "potentiaali pienenee".
T�lt� kohdin on eritt�in vaikeaa n�hd� eroa evoluutikkojen n�kemykseen ; heist� luonnonvalinta valikoi hyv�t ja huonot ominaisuudet ja ett� "jossain on potentiaalisessa muodossa tieto siit� millaista informaatiota eli�iss� voi olla". -Barry Hallin lis�ksi mm. Richard Dawkins on kirjoittanut luonnonvalinnan tuottamasta ennustettavuudesta- Evoluutikoista universumin sis�lt�m�t luonnonlait sek� n�iden keskin�inen vaikutus ymp�rist�ss� sis�lt�v�t "potentiaalisessa muodossa" tiedon "mink�lainen eli� on siihen sopiva ja mik� v�hemm�n sopiva". Spetner tulkitsee osan informaatiosta "nukkuviksi geeneiksi", mutta ei rajaa t�t� ainoaksi vaihtoehdoksi. Sill� miss� "potentiaali -informaation" varasto on, ei ole merkitt�v� asia. "Mikroevoluution potentiaali" voidaan siis varsin hyvin ymm�rt�� my�s "teistiseksi evoluutioksi" eik� oikein mitenk��n "evoluutiokritiikiksi".
Toisin sanoen Spetner ei vetoa ulkoisiin rakenteisiin. H�nen n�kemyksens� kannalta on t�ysin mahdollista, ett� koko eli�kunta olisi l�ht�isin esimerkiksi pienen bakteerin "potentiaalista". T�m� tarkoittaa sit�, ett� Spetner voi ajatella nykyisest� fossiilirekordista sen, ett� "potentiaali -informaatio" selv�sti ajan kanssa muuttuu "aktuaaliseksi informaatioksi".
T�ll� n�kemyksell� on muutamia heikkouksia:
*Ensinn�kin mink�lainen tahansa fossiiliaineisto sopii Spetnerin n�kemykseen, eli fossiileilla n�kemyst� ei voi falsifioida.
*Toisaalta t�m� johtaa mu�s siihen, ett� "rappeutuneiden rakenteiden" esittely ei my�sk��n ole mik��n todiste : Informaatiom��r� kun ei ole pelk�st��n n�kyviss� rakenteissa.
*Kolmanneksi jos potentiaali olisi koodattuna perim��n, ja se vain "nousisi sielt� ajan mittaan esiin", pit�isi mallin selitt�� miten; Ongelmana kun on se, ett� eitoiminnallinen, piilossa oleva, potentiaali on kuitenkin altis satunnaisille mutaatioille, jonka pit�isi johtaa siihen ett� ilman valintapainetta potentiaali tuhoutuu pitk�ll� t�ht�imell�: Toisin sanoen siin� miss� evoluutio EI v�it� ett� ominaisuudet nousisivat esiin sattumalta, vaan p�invastoin korostavat luonnonvalinnan merkityst� siin� ett� toiminta ei tuhoudu, koodattuun, eitoiminnalliseen, potentiaaliin uskova joutuisi hyv�ksym��n "sattumalta esiin nousevan informaation". Potentiaali voisi toki nousta esiin "eisatunnaisin mutaatioin", mutta t�ll�isest� manipulaatiosta ei ole havaintoja.
Spetnerin malli koskee pelk�st��n entsyymeit�, eik� siksi ole sovellettavissa rakenteisiin tai mihink��n muuhun. Spetnerin mallin falsifiointi on kyseenalaista, koska periaatteessa mik� tahansa informaatio voi olla per�isin "jostakin tuntemattomasta potentiaalista" ja vaikka jokin mutaatio muuttaisi jotain entsyymi� "spesifisemm�ksi", on se ainoastaan "spesifisemm�ksi suhteessa tiet�miimme substraatteihin", jolloin ei voida tiet�� onko kyse "aidosta spesifioitumisesta" eli "spesifisyydest� suhteessa kaikkiin substraatteihin". Jos potentiaali katsotaan rajoitetuksi pelk�st��n perim��n, niin tilanne muuttuu. Spetner muuttuu empiriaksi heti, kun tapa mitata potentiaalin suuruus keksit��n, ja heti, kun Spetner onnistuu keksim��n sen mekanismin, joka takaa sen, ett� mutaatiot ovatkin hy�dyllisi�, informaatiota "potentiaalista" esiinkaivavia, sen sijaan ett� olisivat "satunnaisia".
Mutaatiot - uuden rakentaminen vai varastostahaku
"I agree with him that these mutations add information to the B-cell genome."
(Lee Spetner, "Lee Spetner/Edward Max Dialogue", 2006)
On havaittu sellaisia "satunnaisia" mutaatioita, jotka lis��v�t geneettist� informaatiota jopa Spetnerin menetelm�ll� mitattuna. Erityisen vaikeaksi Spetnerin mallille on ollut immuunisysteemi, koska vasta -aineet eiv�t ole yhdistettyn� "nukkuviin geeneihin", vaan ensimm�isell� kerralla kun tauti iskee, siihen sopivaa informaatiota ei ole, vaan se on generoitava satunnaisin mutaatioin. Spetner on my�nt�nyt sen, ett� immuunisysteemi lis�� informaatiota B -solun genomiin. Kyse ei siis ole "potentiaali - informaation esiinkaivamisesta", vaan aidosta informaation kasvusta. Eli jotain, jonka piti h�nen omista m��ritelmist��n k�sin olla mahdotonta.
Spetner esitt�� seuraavia syit� sille,miksi t�m� on mahdollista:
*Mutaatiom��r� on suuri : Arviot v�lill� 100 miljoonaa - 10 miljardia kappaletta.
T�m�n p��telm�n Spetner perustelee, sivulla 103, sill� ett� todenn�k�isyys tuottaa informaatiota "evoluutiosattumalta" saadaan, kun kerrotaan jokaisen v�livaiheen todenn�k�isyydet kesken��n : T�m� todenn�k�isyys taas saavutetaan arvaamalla tarpeeksi monta kertaa. T�ss� n�kemyksess� kummallista on:
-Luonnonvalintaa ei oteta huomioon lainkaan, jolloin v�livaiheiden k�ytt� itse asiassa menett�� merkityksens�. V�livaiheella voi olla merkityst� vain luonnonvalinnan kautta. "V�livaiheilla" luodaan ainoastaan valheellinen mielikuva "evoluution huomioon ottamisessa".
-Miten t�t� selityst� soveltaen informaatiota ei synny luonnossa "miljoonissa vuosissa", kun todenn�k�isyydet pysyv�t ja mutaatiot tuottavat jatkuvasti "arvauksia"? Eik� sen pit�isi olla pikemminkin v�ist�m�t�nt�?
On joka tapauksessa eritt�in erikoista, ett� Spetner, jotka kiist�� "aidosti hy�dyllisten mutaatioiden" olemassaolonkin kaikesta huolimatta itse tarjoaa mutaatioita "tarpeeksi isoissa m��riss� esiintyess��n" selityksen� "aidosti uuden informaation" synnylle. Eih�n "puhtaan todenn�k�isyyden" t�yttymiselle, sellaisenahan Spetner tilanteen laskee- ole ratkaisevaa se, kuinka lyhyess� ajassa yritykset tapahtuvat, vaan siin� kuinka monta kertaa n�in tapahtuu.
*Immuunisysteemiss� k�sitelt�v� informaatiom��r� on pieni
T�m� on "virhekatastrofirajaan" sovellettuna aivan j�rkev� argumentti. Suuret mutaatiom��r�t kuitenkin toimisivat hajottavana voimana. Perim�ss�mme mutaatiom��r� on selv�sti immuunisysteemi� pienempi informaatiom��r� ovat selv�sti suurempi. N�in suurempi informaatiom��r� joudutaan kompensoimaan, eik� virhekatastrofiin ajauduta.
*Systeemi on hienos��detty.
On aivan totta, ett� informaatio "on biteiss�": sek� informaation kasvu ett� sen v�heneminen ovat muutoksia niiss� - luokiteltiin informaatio mill� tavoin tahansa. N�in ollen pelk�t muutokset eiv�t riit�, vaan tarvitaan jotain, joka "tarkastaa" mitk� ovat hyvi� ja mitk� huonoja muutoksia. Immuunisysteemiss� on tapoja tunnistaa hyv�t ja huonot muutokset, Spetner kutstuu t�t� "hienos��d�ksi". Luonnossa vastaavaa ei kutsuta "hienos��d�ksi" vaan "luonnonvalinnaksi" - ja t�t� evoluutikot pit�v�t kovasti arvossa.
*Muutokset eiv�t tallennu perim��n.
Spetnerin k�sitys muuttuu t�ll� perusteella hieman kummalliseksi, koska h�nen v�itteens� on ymm�rrett�viss� "Informaatiota ei voi synty�, jos lopputulos tallennetaan." Lis�ongelmia t�lle tuo se, ett� vaikka onkin totta, ett� immuunisysteemi ei tallenna muutoksia ituradan perim��n, niin silti tiedot tallentuvat immuunisysteemin soluihin: syntyy resistenssi ja sama tauti paranee seuraavalla kerralla nopeammin. Ilmeisesti pelkk� tallennuskaan ei siis voi olla este, vaan ilmeisesti jokin vaikutus on sill� minne se tallennus tehd��n.. Tai sitten t�m� argumentti on n�enn�inen.
Kun n�m� kaikki kohdat otetaan huomioon on helpohko huomata, ett� immuunisysteemi on "analoginen" evoluutioprosesseihin. Evoluutiokin on hienostunut systeemi, jossa ymp�rist� asettaa rajaukset informaatiolle, ja jossa esiintyy runsaasti mutaatioita ja jossa on valintamekanismi.
Yhteenvetoa
"The NREH, on the other hand, postulates nonrandom variation. It does not fulfill the neo-Darwinian agenda in that it does not contribute to a natural explanation of the origin of life."
(Lee Spetner, "Not by Chance!", 1998, sivu 210)
Spetnerin keskittyy evoluutiokritiikkiin, mutta muista poiketen esitt�� my�s vastahypoteesin, "NREH":n.
*Evoluutiohypoteesissaan Spetner nojaa omaan informaatiom��rittelyyns�, joka ei ole kovin osuva -Spetner ei esimerkiksi k�sit�, ett� entsyymien ja substraattien sitoutumisella ei ole "kaikki tai ei mit��n" -luonnetta. H�n my�s laittaa yht�l�isyysmerkit "puhtaan Sattuman" ja "evoluution" v�liin. Spetnerin k�ytt�m� "hienos��t�perustelu", joka sallii informaation kasvun taas on ominaisuuksiltaan kovasti samanlainen, kuin luonnonvalinta.
*NREH:ssa keskeisint� on "evoluution ennalta ohjelmoitu potentiaali", joka m��r�� miten kehitys menee ja mink�laista informaatiota tulee. Spetner ei kuitenkaan selit� miten mutaatiot kaivavat potentiaalista informaatiota, eik� sit� miten t�t� potentiaalia voitaisiin tutkia. H�n paljastaa, ett� osa on "nukkuvia geenej�", mutta t�m� ei ole ainut menetelm�. Spetner ei my�sk��n kerro, syntyyk� "NREH":n piiriss� uutta informaatiota vai ei. Emme loppupeleiss� tied� mik� t�m� "potentiaali" on, emmek� my�sk��n mitk� ovat "NREH":n rajat, ja t�t� kautta kautta meill� ei ole keinoa falsifioida Spetnerin teoriaa: Esimerkiksi havainnot, joissa Spetnerin m��ritelm�n mukainen informaatio lis��ntyy ei falsifioi h�nen malliaan, koska ne voidaan tulkita "tuntemattomasta potentiaalista" johtuviksi. Spetner ei my�sk��n sovella evoluutioon soveltamaansa "Voisiko-se-toimia?" -kaavaansa omaan informaatiomalliinsa: l�hinn� koska h�nen mallinsa ei ainakaan toistaiseksi kerro mit��n sellaista mit� evoluutiokaan ei kertoisi - h�nen mallinsa keskeisimm�t, eli "mutaatioita ohjaava voima" ja "mikroevoluution potentiaali" kun ovat molemmat paitsi tuntemattomia my�s tuntemattoman suuruisia.
Kaikesta huolimatta Spetnerin malli ansaitsee erikoishuomion siit�, ett� se on toistaiseksi ainut kreationistien informaatiomalli, joka on sek� formalisaatio ett� vaihtoehtoinen malli.
Lis�tietoja:
Timo Tiainen,"Kriittinen arvio Spetnerin n�kemyksist�",2004
Ian Musgrave,"Information Theory and Creationism - Spetner and Biological Information",2005
Gert Korthof,"Could it work?",2003
Mark Chu-Carroll, "Using Bad Math to Create Bad Models to Produce Bad Results",2007
"When I say a variation is not random, I mean that the chance of it occurring has something to do with how the organism adapts to its environment"
(Lee Spetner, "Not By Chance!", 1998, sivu 175)
Spetnerin m��rittelyn pohjalta luonnossa ilmenev�n informaation mittaaminen on mahdotonta, koska se vaatisi sen, ett� tutkittavan entsyymin toiminta tulisi testata suhteessa joka ikiseen universumissa mahdolliseen substraattiin : Ja n�iden lukum��r�� saati rakennetta emme edes tied�, saati sitten saa yhteiseen tilaan entsyymin kanssa.
Spetnerin informaatiomallissa oletetaan,ett� geeni toimii "joko-tai" mekanismilla ilman v�livaiheita; t�m�n ns. geeniperheet osoittavat v��r�ksi : niiss� useat samankaltaiset geenit hoitavat erilaisia, joskus osittain limitt�isi�kin, toimintoja. (Lokki, Saura & Tigerstedt,"Evoluutio ja populaatiot.",1986) N�inollen toiminto voi muuttua ja kun kahdentuminen on mahdollista, voi kahdentuma + toisen muutos johtaa uuden informaation syntyyn "eritt�in mikroevolutiivisesti".
Spetnerin informaatiom��rittelyss� on my�skin joitain puutteita. Siin� ei esimerkiksi oteta lainkaan huomioon toimintalaatua. Jos entsyymi toimii mitenk��n on se informaatiota; Spetnerin informaatio on "joko tai", vaikka entsyymien toiminta ei t�ll�ist� ole, ja eli�n toiminnallisuuden kannalta on kuitenkin suunnatonta merkityst� sill� mill� teholla entsyymi toimii."Entsyymi - substraattiyhdistym� muodostuu heikkojen sidosten, kuten vetysidosten ja ionisidosten kautta. Yhtym� muodostuu sit� helpommin mit� enemm�n n�it� sidoksia on; toisaalta sidoksia on sit� enemm�n mit� paremmin entsyymin aktiivinen kohta ja substraatti sopivat toisiinsa. Entsyymin valikoivuus selittyy t�t� taustaa vasten. Yksi substraatti sopii entsyymiin parhaiten, muut huonommin."
(Jyrki Heino, Matti Vuento, "Solubiologia", sivu 61) My�sk��n sidoksen lujuutta Spetnerin mallissa ei k�sitell�.
Spetnerin kaavan mukaan informaatiom��r� kasvaa sit� mukaa, mit� enemm�n on olemassa substraatteja, joihin entsyymi ei vaikuta, joten uusia substraatteja synnytt�m�ll� informaatiom��r� voisi kasvaa: T�m� johtaa my�s siihen ett� sin� hetken� kun entsyymi menett�� kokonaan toimintansa, sill� ei ole substraattia ja entsyymin informaatio nousee "Spetnerin tasolla valtavaksi" koska substraattien m��r� joihin entsyymi ei vaikuta on "kaikki mahdolliset". T�lt� kannalta eli�, jonka sis�ll� ei olisi ainuttakaan toimivaa entsyymi� sis�lt�isi "maksimaalisesti informaatiota". -Joskin juuri t�m� ominaisuus selitt�� miten "nukkuva geeni" voi informaatiota kadottamalla muuttua toiminnalliseksi. Ennusteena Spetnerin teoriassa on, ett� satunnaiset mutaatiot tuhoaisivat informaatiota, jolloin satunnaismutaatiory�ppy aina kasvattaisi "entsyymien laaja -alaisuutta" - jolloin koetta jatkettaessa lopulta jokainen entsyymi toimisi jokaisen substraatin kanssa.
Spetnerin tunnetuimmat s��nn�t ovat "satunnaiset muutokset eiv�t tuota informaatiota" ja ett� "mutaatiot karsivat informaatiota"
[] => v [] a // satunnaisia muutoksia, m��r�isyys ei kasva.
a valikoituu // v + [] katoavat, informaatio v�henee
a => a ai ab // satunnaisia muutoksia, m��r�isyys ei kasva
ab => valikoituu // informaatio v�henee
...
a...y => a...z, a..ya //satunnaisia muutoksia, m��r�isyys ei kasva
a...z => valikoituu // informaatio v�henee
Spetnerlogiikalla informaation m��r� on koko ajan v�hentynyt, vaikka se on kasvanut nollasta 123:n bittiin ja informaatio on eksaktia. Salakuljetettua informaatiota siin� on samassa suhteessa miss� luonnossakin. Algoritmi tiet�� ainoastaan kuinka hyv� satunnaisten mutaatioiden muuntelema ratkaisu on tiettyjen kriteereiden valossa(mahdollisimman pitk� aakkosj�rjestys). Samoin luonnossa ymp�r�iv�t luonnonlait m��r��v�t sen mitk� muodot s�ilyv�t ja mitk� eiv�t.
Evoluution potentiaali
"A classic example is to remove the gene that allows E. coli bacteria to digest lactose, but then give the colony of bacteria only lactose to eat. Of the millions in a colony, some bacteria re-evolve the ability to hydrolyze lactose and survive."
(Science Daily Barry G. Hallin tutkimuksesta, "Darwin's Time Machine: Scientists Begin Predicting Evolution's Next Step", 20.Maal.2002.)
Informaationtuotto -ongelmien varalle Spetner on kehitt�nyt "mikroevoluution potentiaaliin" eli "esiohjelmoitu kehitys". T�ss� "eli�t ovat hienos��detyt", jolloin ilmestyy "yll�tt�v��kin muuntelua".
H�n vetoaa usein Barry G. Hallin 1982 tekem��n tutkimukseen. Siin� osoitettiin, ett� populaatioon ilmestyy s��nn�nmukaisesti ja sangen nopeasti tietyt muutokset tietyss� valintapaineessa. Spetnerin tulkinta kyseisest� tutkimuksesta on, ett� hy�dylliset mutaatiot ovat liian yleisi�, jolloin ne eiv�t ole "aidosti informaatiota luovia"(~Spetnerin "truly beneficial") vaan kaivavat informaation "potentiaalistaan".
Barry Hall on suorittanut sittemmin t�ydent�vi� tutkimuksia, mutta n�ist� Spetner ei puhu. Onhan se toisaalta ik�v��, ett� jo "klassisessa esimerkiss�" ensin suoritetaan "deleetio", eli koko ominaisuuden poisto ja se silti kehittyy takaisin - suorastaan ennustettavan pilkuntarkasti. "Potentiaali" ei siis voi olla perim�ss�. Vaikuttaa "evoluution hyvyysfunktioilta" eli tutummin "luonnonvalinnalta": T�ll�in Spetnerin "mikroevoluution potentiaali" tarkoittaa yksinkertaisesti samaa asiaa, mihin viittaa my�s Ernst Meyer 2003 julkaistun kirjansa "Evoluutio" sivulla 157 lauseessa "Koska kaikki kuviteltavissa olevat hy�dylliset mutaatiot ovat jo valikoituneet vakaissa elinymp�rist�iss�, uusien hy�dyllisten mutaatioiden syntyminen on melko harvinaista." Eli evoluutiolla on olemassa (lokaali)optimi, jonka "yli tai ohi" informaatiota ei voi synty�. K�yt�nn�ss� Spetnerin "neodarwinismin kritiikki" lakkaa olemasta "neodarwinismin kritiikki�", jos h�n my�nt�� luonnonvalinnan t�m�n ominaisuuden, joten ainut vaihtoehto on vetoaminen "nukkuviin geeneihin". Eli pelkk� klassinen "sokerijuurikkaan sokeripitoisuusesimerkki" tai muut vastaavat, jossa osoitetaan, ett� "informaatiolla on lokaali optimi" ei riit�: Ollakseen "evoluutiolle ongelma" t�ytyy my�s pysty� erottamaan "lokaali optimi" ja "mikroevoluution koko kyky" toisistaan ja osoittamaan, ett� kyse ei ole pelk�st� "lokaalista optimista": Lokaali optimi -v�ite kun on ongelmallinen siksi ett� vaikka evoluutioalgoritmeissa voi lokaali optimi olla pysyv�, luonnossa "hyvyysfunktiot" muuttuvat ajan kanssa. Jopa hyvin yksinkertainen esimerkki osoittaa sen ett� hyvyysfunktiot ja "lokaalit optimit" evoluutiossa ovat pakotettuja muuttumaan, ne eiv�t ole mik��n sein� kuin korkeintaan v�liaikaisesti:
Oletetaan nyt ett� meill� on hyvyysfunktio jossa on lokaali optimi, joka ei kuitenkaan ole "korkein mahdollinen huippu". Sitten l�yd�mme organismin, joka kiipe�� siihen. Kun evoluutio saavuttaa maksimin, evoluutio lakkaa etenem�st�, jonka seurauksena juuri sen lokaalin optimin kohdalla alkaa olemaan ruuhkaa: Yksil�t kuluttavat samaa ravintoa, joka taas vaikuttaa hyvyysfunktioon. Seurauksena lokaalin optimin paikka muuttuu.
Lis�ksi evoluutio on moniulotteinen, joten pit�isi todistaa ett� optimi olisi rajattu joka suuntaan. Jos meill� on hyvyysavaruus, jolla on X ja Y akseli, ei voida sanoa ett� se ett� kohde on sellaisella kohdalla ett� sen Y on lokaalissa optimissa, se ei tarkoita ett� se ei voisi muuttua paremmaksi, se voi liikkua X:n kautta, ja t�m� X:n muutos voi jopa tarjota uuden kohdan, jossa Y ei en�� olekaan sidottu. (Eli lukkiutumia voi synty� ja ne voivat vaihtaa paikkaa.) "Lokaali optimi" ei siksi toimi "mikroevoluutioesteen� jota ei voi ylitt��".
Eli "hyvyysfunktiot" ovat tuskin koskaan staattisia ja "useampi ulottuvuus" tuo lis�ksi uusia kiertoteit�.
Toisin sanoen, Spetnerist� s��nn�nmukainen kehitys ei suinkaan osoita, ett� h�nen omat laskelmansa "rappeutumisen pakosta" ja "evoluution satunnaisuudesta eli kyvytt�myydest� tuottaa ennusteita", olisivat v��r�ss�. H�n ei my�sk��n p���ttele, ett� "informaation trendi on nouseva", vaan sen sijaan se kertoo siit�, ett� muuntelumahdollisuuksien m��r� on rajallinen ja n�ist� vaihtoehdoista valikoituu tietty "sattumanvaraisesti eli ymp�rist�st� riippuen". Samanaikaisesti, kun "aktuaalinen, havaittava informaatio" kasvaa, niin "potentiaali pienenee".
T�lt� kohdin on eritt�in vaikeaa n�hd� eroa evoluutikkojen n�kemykseen ; heist� luonnonvalinta valikoi hyv�t ja huonot ominaisuudet ja ett� "jossain on potentiaalisessa muodossa tieto siit� millaista informaatiota eli�iss� voi olla". -Barry Hallin lis�ksi mm. Richard Dawkins on kirjoittanut luonnonvalinnan tuottamasta ennustettavuudesta- Evoluutikoista universumin sis�lt�m�t luonnonlait sek� n�iden keskin�inen vaikutus ymp�rist�ss� sis�lt�v�t "potentiaalisessa muodossa" tiedon "mink�lainen eli� on siihen sopiva ja mik� v�hemm�n sopiva". Spetner tulkitsee osan informaatiosta "nukkuviksi geeneiksi", mutta ei rajaa t�t� ainoaksi vaihtoehdoksi. Sill� miss� "potentiaali -informaation" varasto on, ei ole merkitt�v� asia. "Mikroevoluution potentiaali" voidaan siis varsin hyvin ymm�rt�� my�s "teistiseksi evoluutioksi" eik� oikein mitenk��n "evoluutiokritiikiksi".
Toisin sanoen Spetner ei vetoa ulkoisiin rakenteisiin. H�nen n�kemyksens� kannalta on t�ysin mahdollista, ett� koko eli�kunta olisi l�ht�isin esimerkiksi pienen bakteerin "potentiaalista". T�m� tarkoittaa sit�, ett� Spetner voi ajatella nykyisest� fossiilirekordista sen, ett� "potentiaali -informaatio" selv�sti ajan kanssa muuttuu "aktuaaliseksi informaatioksi".
T�ll� n�kemyksell� on muutamia heikkouksia:
*Ensinn�kin mink�lainen tahansa fossiiliaineisto sopii Spetnerin n�kemykseen, eli fossiileilla n�kemyst� ei voi falsifioida.
*Toisaalta t�m� johtaa mu�s siihen, ett� "rappeutuneiden rakenteiden" esittely ei my�sk��n ole mik��n todiste : Informaatiom��r� kun ei ole pelk�st��n n�kyviss� rakenteissa.
*Kolmanneksi jos potentiaali olisi koodattuna perim��n, ja se vain "nousisi sielt� ajan mittaan esiin", pit�isi mallin selitt�� miten; Ongelmana kun on se, ett� eitoiminnallinen, piilossa oleva, potentiaali on kuitenkin altis satunnaisille mutaatioille, jonka pit�isi johtaa siihen ett� ilman valintapainetta potentiaali tuhoutuu pitk�ll� t�ht�imell�: Toisin sanoen siin� miss� evoluutio EI v�it� ett� ominaisuudet nousisivat esiin sattumalta, vaan p�invastoin korostavat luonnonvalinnan merkityst� siin� ett� toiminta ei tuhoudu, koodattuun, eitoiminnalliseen, potentiaaliin uskova joutuisi hyv�ksym��n "sattumalta esiin nousevan informaation". Potentiaali voisi toki nousta esiin "eisatunnaisin mutaatioin", mutta t�ll�isest� manipulaatiosta ei ole havaintoja.
Spetnerin malli koskee pelk�st��n entsyymeit�, eik� siksi ole sovellettavissa rakenteisiin tai mihink��n muuhun. Spetnerin mallin falsifiointi on kyseenalaista, koska periaatteessa mik� tahansa informaatio voi olla per�isin "jostakin tuntemattomasta potentiaalista" ja vaikka jokin mutaatio muuttaisi jotain entsyymi� "spesifisemm�ksi", on se ainoastaan "spesifisemm�ksi suhteessa tiet�miimme substraatteihin", jolloin ei voida tiet�� onko kyse "aidosta spesifioitumisesta" eli "spesifisyydest� suhteessa kaikkiin substraatteihin". Jos potentiaali katsotaan rajoitetuksi pelk�st��n perim��n, niin tilanne muuttuu. Spetner muuttuu empiriaksi heti, kun tapa mitata potentiaalin suuruus keksit��n, ja heti, kun Spetner onnistuu keksim��n sen mekanismin, joka takaa sen, ett� mutaatiot ovatkin hy�dyllisi�, informaatiota "potentiaalista" esiinkaivavia, sen sijaan ett� olisivat "satunnaisia".
Mutaatiot - uuden rakentaminen vai varastostahaku
"I agree with him that these mutations add information to the B-cell genome."
(Lee Spetner, "Lee Spetner/Edward Max Dialogue", 2006)
On havaittu sellaisia "satunnaisia" mutaatioita, jotka lis��v�t geneettist� informaatiota jopa Spetnerin menetelm�ll� mitattuna. Erityisen vaikeaksi Spetnerin mallille on ollut immuunisysteemi, koska vasta -aineet eiv�t ole yhdistettyn� "nukkuviin geeneihin", vaan ensimm�isell� kerralla kun tauti iskee, siihen sopivaa informaatiota ei ole, vaan se on generoitava satunnaisin mutaatioin. Spetner on my�nt�nyt sen, ett� immuunisysteemi lis�� informaatiota B -solun genomiin. Kyse ei siis ole "potentiaali - informaation esiinkaivamisesta", vaan aidosta informaation kasvusta. Eli jotain, jonka piti h�nen omista m��ritelmist��n k�sin olla mahdotonta.
Spetner esitt�� seuraavia syit� sille,miksi t�m� on mahdollista:
*Mutaatiom��r� on suuri : Arviot v�lill� 100 miljoonaa - 10 miljardia kappaletta.
T�m�n p��telm�n Spetner perustelee, sivulla 103, sill� ett� todenn�k�isyys tuottaa informaatiota "evoluutiosattumalta" saadaan, kun kerrotaan jokaisen v�livaiheen todenn�k�isyydet kesken��n : T�m� todenn�k�isyys taas saavutetaan arvaamalla tarpeeksi monta kertaa. T�ss� n�kemyksess� kummallista on:
-Luonnonvalintaa ei oteta huomioon lainkaan, jolloin v�livaiheiden k�ytt� itse asiassa menett�� merkityksens�. V�livaiheella voi olla merkityst� vain luonnonvalinnan kautta. "V�livaiheilla" luodaan ainoastaan valheellinen mielikuva "evoluution huomioon ottamisessa".
-Miten t�t� selityst� soveltaen informaatiota ei synny luonnossa "miljoonissa vuosissa", kun todenn�k�isyydet pysyv�t ja mutaatiot tuottavat jatkuvasti "arvauksia"? Eik� sen pit�isi olla pikemminkin v�ist�m�t�nt�?
On joka tapauksessa eritt�in erikoista, ett� Spetner, jotka kiist�� "aidosti hy�dyllisten mutaatioiden" olemassaolonkin kaikesta huolimatta itse tarjoaa mutaatioita "tarpeeksi isoissa m��riss� esiintyess��n" selityksen� "aidosti uuden informaation" synnylle. Eih�n "puhtaan todenn�k�isyyden" t�yttymiselle, sellaisenahan Spetner tilanteen laskee- ole ratkaisevaa se, kuinka lyhyess� ajassa yritykset tapahtuvat, vaan siin� kuinka monta kertaa n�in tapahtuu.
*Immuunisysteemiss� k�sitelt�v� informaatiom��r� on pieni
T�m� on "virhekatastrofirajaan" sovellettuna aivan j�rkev� argumentti. Suuret mutaatiom��r�t kuitenkin toimisivat hajottavana voimana. Perim�ss�mme mutaatiom��r� on selv�sti immuunisysteemi� pienempi informaatiom��r� ovat selv�sti suurempi. N�in suurempi informaatiom��r� joudutaan kompensoimaan, eik� virhekatastrofiin ajauduta.
*Systeemi on hienos��detty.
On aivan totta, ett� informaatio "on biteiss�": sek� informaation kasvu ett� sen v�heneminen ovat muutoksia niiss� - luokiteltiin informaatio mill� tavoin tahansa. N�in ollen pelk�t muutokset eiv�t riit�, vaan tarvitaan jotain, joka "tarkastaa" mitk� ovat hyvi� ja mitk� huonoja muutoksia. Immuunisysteemiss� on tapoja tunnistaa hyv�t ja huonot muutokset, Spetner kutstuu t�t� "hienos��d�ksi". Luonnossa vastaavaa ei kutsuta "hienos��d�ksi" vaan "luonnonvalinnaksi" - ja t�t� evoluutikot pit�v�t kovasti arvossa.
*Muutokset eiv�t tallennu perim��n.
Spetnerin k�sitys muuttuu t�ll� perusteella hieman kummalliseksi, koska h�nen v�itteens� on ymm�rrett�viss� "Informaatiota ei voi synty�, jos lopputulos tallennetaan." Lis�ongelmia t�lle tuo se, ett� vaikka onkin totta, ett� immuunisysteemi ei tallenna muutoksia ituradan perim��n, niin silti tiedot tallentuvat immuunisysteemin soluihin: syntyy resistenssi ja sama tauti paranee seuraavalla kerralla nopeammin. Ilmeisesti pelkk� tallennuskaan ei siis voi olla este, vaan ilmeisesti jokin vaikutus on sill� minne se tallennus tehd��n.. Tai sitten t�m� argumentti on n�enn�inen.
Kun n�m� kaikki kohdat otetaan huomioon on helpohko huomata, ett� immuunisysteemi on "analoginen" evoluutioprosesseihin. Evoluutiokin on hienostunut systeemi, jossa ymp�rist� asettaa rajaukset informaatiolle, ja jossa esiintyy runsaasti mutaatioita ja jossa on valintamekanismi.
Yhteenvetoa
"The NREH, on the other hand, postulates nonrandom variation. It does not fulfill the neo-Darwinian agenda in that it does not contribute to a natural explanation of the origin of life."
(Lee Spetner, "Not by Chance!", 1998, sivu 210)
Spetnerin keskittyy evoluutiokritiikkiin, mutta muista poiketen esitt�� my�s vastahypoteesin, "NREH":n.
*Evoluutiohypoteesissaan Spetner nojaa omaan informaatiom��rittelyyns�, joka ei ole kovin osuva -Spetner ei esimerkiksi k�sit�, ett� entsyymien ja substraattien sitoutumisella ei ole "kaikki tai ei mit��n" -luonnetta. H�n my�s laittaa yht�l�isyysmerkit "puhtaan Sattuman" ja "evoluution" v�liin. Spetnerin k�ytt�m� "hienos��t�perustelu", joka sallii informaation kasvun taas on ominaisuuksiltaan kovasti samanlainen, kuin luonnonvalinta.
*NREH:ssa keskeisint� on "evoluution ennalta ohjelmoitu potentiaali", joka m��r�� miten kehitys menee ja mink�laista informaatiota tulee. Spetner ei kuitenkaan selit� miten mutaatiot kaivavat potentiaalista informaatiota, eik� sit� miten t�t� potentiaalia voitaisiin tutkia. H�n paljastaa, ett� osa on "nukkuvia geenej�", mutta t�m� ei ole ainut menetelm�. Spetner ei my�sk��n kerro, syntyyk� "NREH":n piiriss� uutta informaatiota vai ei. Emme loppupeleiss� tied� mik� t�m� "potentiaali" on, emmek� my�sk��n mitk� ovat "NREH":n rajat, ja t�t� kautta kautta meill� ei ole keinoa falsifioida Spetnerin teoriaa: Esimerkiksi havainnot, joissa Spetnerin m��ritelm�n mukainen informaatio lis��ntyy ei falsifioi h�nen malliaan, koska ne voidaan tulkita "tuntemattomasta potentiaalista" johtuviksi. Spetner ei my�sk��n sovella evoluutioon soveltamaansa "Voisiko-se-toimia?" -kaavaansa omaan informaatiomalliinsa: l�hinn� koska h�nen mallinsa ei ainakaan toistaiseksi kerro mit��n sellaista mit� evoluutiokaan ei kertoisi - h�nen mallinsa keskeisimm�t, eli "mutaatioita ohjaava voima" ja "mikroevoluution potentiaali" kun ovat molemmat paitsi tuntemattomia my�s tuntemattoman suuruisia.
Kaikesta huolimatta Spetnerin malli ansaitsee erikoishuomion siit�, ett� se on toistaiseksi ainut kreationistien informaatiomalli, joka on sek� formalisaatio ett� vaihtoehtoinen malli.
Lis�tietoja:
Timo Tiainen,"Kriittinen arvio Spetnerin n�kemyksist�",2004
Ian Musgrave,"Information Theory and Creationism - Spetner and Biological Information",2005
Gert Korthof,"Could it work?",2003
Mark Chu-Carroll, "Using Bad Math to Create Bad Models to Produce Bad Results",2007
Spetnerin informaatiomallissa oletetaan,ett� geeni toimii "joko-tai" mekanismilla ilman v�livaiheita; t�m�n ns. geeniperheet osoittavat v��r�ksi : niiss� useat samankaltaiset geenit hoitavat erilaisia, joskus osittain limitt�isi�kin, toimintoja. (Lokki, Saura & Tigerstedt,"Evoluutio ja populaatiot.",1986) N�inollen toiminto voi muuttua ja kun kahdentuminen on mahdollista, voi kahdentuma + toisen muutos johtaa uuden informaation syntyyn "eritt�in mikroevolutiivisesti".
Spetnerin informaatiom��rittelyss� on my�skin joitain puutteita. Siin� ei esimerkiksi oteta lainkaan huomioon toimintalaatua. Jos entsyymi toimii mitenk��n on se informaatiota; Spetnerin informaatio on "joko tai", vaikka entsyymien toiminta ei t�ll�ist� ole, ja eli�n toiminnallisuuden kannalta on kuitenkin suunnatonta merkityst� sill� mill� teholla entsyymi toimii.
"Entsyymi - substraattiyhdistym� muodostuu heikkojen sidosten, kuten vetysidosten ja ionisidosten kautta. Yhtym� muodostuu sit� helpommin mit� enemm�n n�it� sidoksia on; toisaalta sidoksia on sit� enemm�n mit� paremmin entsyymin aktiivinen kohta ja substraatti sopivat toisiinsa. Entsyymin valikoivuus selittyy t�t� taustaa vasten. Yksi substraatti sopii entsyymiin parhaiten, muut huonommin."
(Jyrki Heino, Matti Vuento, "Solubiologia", sivu 61) My�sk��n sidoksen lujuutta Spetnerin mallissa ei k�sitell�.
Spetnerin kaavan mukaan informaatiom��r� kasvaa sit� mukaa, mit� enemm�n on olemassa substraatteja, joihin entsyymi ei vaikuta, joten uusia substraatteja synnytt�m�ll� informaatiom��r� voisi kasvaa: T�m� johtaa my�s siihen ett� sin� hetken� kun entsyymi menett�� kokonaan toimintansa, sill� ei ole substraattia ja entsyymin informaatio nousee "Spetnerin tasolla valtavaksi" koska substraattien m��r� joihin entsyymi ei vaikuta on "kaikki mahdolliset". T�lt� kannalta eli�, jonka sis�ll� ei olisi ainuttakaan toimivaa entsyymi� sis�lt�isi "maksimaalisesti informaatiota". -Joskin juuri t�m� ominaisuus selitt�� miten "nukkuva geeni" voi informaatiota kadottamalla muuttua toiminnalliseksi. Ennusteena Spetnerin teoriassa on, ett� satunnaiset mutaatiot tuhoaisivat informaatiota, jolloin satunnaismutaatiory�ppy aina kasvattaisi "entsyymien laaja -alaisuutta" - jolloin koetta jatkettaessa lopulta jokainen entsyymi toimisi jokaisen substraatin kanssa.
Spetnerin tunnetuimmat s��nn�t ovat "satunnaiset muutokset eiv�t tuota informaatiota" ja ett� "mutaatiot karsivat informaatiota"
[] => v [] a // satunnaisia muutoksia, m��r�isyys ei kasva.
a valikoituu // v + [] katoavat, informaatio v�henee
a => a ai ab // satunnaisia muutoksia, m��r�isyys ei kasva
ab => valikoituu // informaatio v�henee
...
a...y => a...z, a..ya //satunnaisia muutoksia, m��r�isyys ei kasva
a...z => valikoituu // informaatio v�henee
Spetnerlogiikalla informaation m��r� on koko ajan v�hentynyt, vaikka se on kasvanut nollasta 123:n bittiin ja informaatio on eksaktia. Salakuljetettua informaatiota siin� on samassa suhteessa miss� luonnossakin. Algoritmi tiet�� ainoastaan kuinka hyv� satunnaisten mutaatioiden muuntelema ratkaisu on tiettyjen kriteereiden valossa(mahdollisimman pitk� aakkosj�rjestys). Samoin luonnossa ymp�r�iv�t luonnonlait m��r��v�t sen mitk� muodot s�ilyv�t ja mitk� eiv�t.
Evoluution potentiaali
"A classic example is to remove the gene that allows E. coli bacteria to digest lactose, but then give the colony of bacteria only lactose to eat. Of the millions in a colony, some bacteria re-evolve the ability to hydrolyze lactose and survive."
(Science Daily Barry G. Hallin tutkimuksesta, "Darwin's Time Machine: Scientists Begin Predicting Evolution's Next Step", 20.Maal.2002.)
Informaationtuotto -ongelmien varalle Spetner on kehitt�nyt "mikroevoluution potentiaaliin" eli "esiohjelmoitu kehitys". T�ss� "eli�t ovat hienos��detyt", jolloin ilmestyy "yll�tt�v��kin muuntelua".
H�n vetoaa usein Barry G. Hallin 1982 tekem��n tutkimukseen. Siin� osoitettiin, ett� populaatioon ilmestyy s��nn�nmukaisesti ja sangen nopeasti tietyt muutokset tietyss� valintapaineessa. Spetnerin tulkinta kyseisest� tutkimuksesta on, ett� hy�dylliset mutaatiot ovat liian yleisi�, jolloin ne eiv�t ole "aidosti informaatiota luovia"(~Spetnerin "truly beneficial") vaan kaivavat informaation "potentiaalistaan".
Barry Hall on suorittanut sittemmin t�ydent�vi� tutkimuksia, mutta n�ist� Spetner ei puhu. Onhan se toisaalta ik�v��, ett� jo "klassisessa esimerkiss�" ensin suoritetaan "deleetio", eli koko ominaisuuden poisto ja se silti kehittyy takaisin - suorastaan ennustettavan pilkuntarkasti. "Potentiaali" ei siis voi olla perim�ss�. Vaikuttaa "evoluution hyvyysfunktioilta" eli tutummin "luonnonvalinnalta": T�ll�in Spetnerin "mikroevoluution potentiaali" tarkoittaa yksinkertaisesti samaa asiaa, mihin viittaa my�s Ernst Meyer 2003 julkaistun kirjansa "Evoluutio" sivulla 157 lauseessa "Koska kaikki kuviteltavissa olevat hy�dylliset mutaatiot ovat jo valikoituneet vakaissa elinymp�rist�iss�, uusien hy�dyllisten mutaatioiden syntyminen on melko harvinaista." Eli evoluutiolla on olemassa (lokaali)optimi, jonka "yli tai ohi" informaatiota ei voi synty�. K�yt�nn�ss� Spetnerin "neodarwinismin kritiikki" lakkaa olemasta "neodarwinismin kritiikki�", jos h�n my�nt�� luonnonvalinnan t�m�n ominaisuuden, joten ainut vaihtoehto on vetoaminen "nukkuviin geeneihin". Eli pelkk� klassinen "sokerijuurikkaan sokeripitoisuusesimerkki" tai muut vastaavat, jossa osoitetaan, ett� "informaatiolla on lokaali optimi" ei riit�: Ollakseen "evoluutiolle ongelma" t�ytyy my�s pysty� erottamaan "lokaali optimi" ja "mikroevoluution koko kyky" toisistaan ja osoittamaan, ett� kyse ei ole pelk�st� "lokaalista optimista": Lokaali optimi -v�ite kun on ongelmallinen siksi ett� vaikka evoluutioalgoritmeissa voi lokaali optimi olla pysyv�, luonnossa "hyvyysfunktiot" muuttuvat ajan kanssa. Jopa hyvin yksinkertainen esimerkki osoittaa sen ett� hyvyysfunktiot ja "lokaalit optimit" evoluutiossa ovat pakotettuja muuttumaan, ne eiv�t ole mik��n sein� kuin korkeintaan v�liaikaisesti:
Oletetaan nyt ett� meill� on hyvyysfunktio jossa on lokaali optimi, joka ei kuitenkaan ole "korkein mahdollinen huippu". Sitten l�yd�mme organismin, joka kiipe�� siihen. Kun evoluutio saavuttaa maksimin, evoluutio lakkaa etenem�st�, jonka seurauksena juuri sen lokaalin optimin kohdalla alkaa olemaan ruuhkaa: Yksil�t kuluttavat samaa ravintoa, joka taas vaikuttaa hyvyysfunktioon. Seurauksena lokaalin optimin paikka muuttuu.
Lis�ksi evoluutio on moniulotteinen, joten pit�isi todistaa ett� optimi olisi rajattu joka suuntaan. Jos meill� on hyvyysavaruus, jolla on X ja Y akseli, ei voida sanoa ett� se ett� kohde on sellaisella kohdalla ett� sen Y on lokaalissa optimissa, se ei tarkoita ett� se ei voisi muuttua paremmaksi, se voi liikkua X:n kautta, ja t�m� X:n muutos voi jopa tarjota uuden kohdan, jossa Y ei en�� olekaan sidottu. (Eli lukkiutumia voi synty� ja ne voivat vaihtaa paikkaa.) "Lokaali optimi" ei siksi toimi "mikroevoluutioesteen� jota ei voi ylitt��".
Eli "hyvyysfunktiot" ovat tuskin koskaan staattisia ja "useampi ulottuvuus" tuo lis�ksi uusia kiertoteit�.
Toisin sanoen, Spetnerist� s��nn�nmukainen kehitys ei suinkaan osoita, ett� h�nen omat laskelmansa "rappeutumisen pakosta" ja "evoluution satunnaisuudesta eli kyvytt�myydest� tuottaa ennusteita", olisivat v��r�ss�. H�n ei my�sk��n p���ttele, ett� "informaation trendi on nouseva", vaan sen sijaan se kertoo siit�, ett� muuntelumahdollisuuksien m��r� on rajallinen ja n�ist� vaihtoehdoista valikoituu tietty "sattumanvaraisesti eli ymp�rist�st� riippuen". Samanaikaisesti, kun "aktuaalinen, havaittava informaatio" kasvaa, niin "potentiaali pienenee".
T�lt� kohdin on eritt�in vaikeaa n�hd� eroa evoluutikkojen n�kemykseen ; heist� luonnonvalinta valikoi hyv�t ja huonot ominaisuudet ja ett� "jossain on potentiaalisessa muodossa tieto siit� millaista informaatiota eli�iss� voi olla". -Barry Hallin lis�ksi mm. Richard Dawkins on kirjoittanut luonnonvalinnan tuottamasta ennustettavuudesta- Evoluutikoista universumin sis�lt�m�t luonnonlait sek� n�iden keskin�inen vaikutus ymp�rist�ss� sis�lt�v�t "potentiaalisessa muodossa" tiedon "mink�lainen eli� on siihen sopiva ja mik� v�hemm�n sopiva". Spetner tulkitsee osan informaatiosta "nukkuviksi geeneiksi", mutta ei rajaa t�t� ainoaksi vaihtoehdoksi. Sill� miss� "potentiaali -informaation" varasto on, ei ole merkitt�v� asia. "Mikroevoluution potentiaali" voidaan siis varsin hyvin ymm�rt�� my�s "teistiseksi evoluutioksi" eik� oikein mitenk��n "evoluutiokritiikiksi".
Toisin sanoen Spetner ei vetoa ulkoisiin rakenteisiin. H�nen n�kemyksens� kannalta on t�ysin mahdollista, ett� koko eli�kunta olisi l�ht�isin esimerkiksi pienen bakteerin "potentiaalista". T�m� tarkoittaa sit�, ett� Spetner voi ajatella nykyisest� fossiilirekordista sen, ett� "potentiaali -informaatio" selv�sti ajan kanssa muuttuu "aktuaaliseksi informaatioksi".
T�ll� n�kemyksell� on muutamia heikkouksia:
*Ensinn�kin mink�lainen tahansa fossiiliaineisto sopii Spetnerin n�kemykseen, eli fossiileilla n�kemyst� ei voi falsifioida.
*Toisaalta t�m� johtaa mu�s siihen, ett� "rappeutuneiden rakenteiden" esittely ei my�sk��n ole mik��n todiste : Informaatiom��r� kun ei ole pelk�st��n n�kyviss� rakenteissa.
*Kolmanneksi jos potentiaali olisi koodattuna perim��n, ja se vain "nousisi sielt� ajan mittaan esiin", pit�isi mallin selitt�� miten; Ongelmana kun on se, ett� eitoiminnallinen, piilossa oleva, potentiaali on kuitenkin altis satunnaisille mutaatioille, jonka pit�isi johtaa siihen ett� ilman valintapainetta potentiaali tuhoutuu pitk�ll� t�ht�imell�: Toisin sanoen siin� miss� evoluutio EI v�it� ett� ominaisuudet nousisivat esiin sattumalta, vaan p�invastoin korostavat luonnonvalinnan merkityst� siin� ett� toiminta ei tuhoudu, koodattuun, eitoiminnalliseen, potentiaaliin uskova joutuisi hyv�ksym��n "sattumalta esiin nousevan informaation". Potentiaali voisi toki nousta esiin "eisatunnaisin mutaatioin", mutta t�ll�isest� manipulaatiosta ei ole havaintoja.
Spetnerin malli koskee pelk�st��n entsyymeit�, eik� siksi ole sovellettavissa rakenteisiin tai mihink��n muuhun. Spetnerin mallin falsifiointi on kyseenalaista, koska periaatteessa mik� tahansa informaatio voi olla per�isin "jostakin tuntemattomasta potentiaalista" ja vaikka jokin mutaatio muuttaisi jotain entsyymi� "spesifisemm�ksi", on se ainoastaan "spesifisemm�ksi suhteessa tiet�miimme substraatteihin", jolloin ei voida tiet�� onko kyse "aidosta spesifioitumisesta" eli "spesifisyydest� suhteessa kaikkiin substraatteihin". Jos potentiaali katsotaan rajoitetuksi pelk�st��n perim��n, niin tilanne muuttuu. Spetner muuttuu empiriaksi heti, kun tapa mitata potentiaalin suuruus keksit��n, ja heti, kun Spetner onnistuu keksim��n sen mekanismin, joka takaa sen, ett� mutaatiot ovatkin hy�dyllisi�, informaatiota "potentiaalista" esiinkaivavia, sen sijaan ett� olisivat "satunnaisia".
Mutaatiot - uuden rakentaminen vai varastostahaku
"I agree with him that these mutations add information to the B-cell genome."
(Lee Spetner, "Lee Spetner/Edward Max Dialogue", 2006)
On havaittu sellaisia "satunnaisia" mutaatioita, jotka lis��v�t geneettist� informaatiota jopa Spetnerin menetelm�ll� mitattuna. Erityisen vaikeaksi Spetnerin mallille on ollut immuunisysteemi, koska vasta -aineet eiv�t ole yhdistettyn� "nukkuviin geeneihin", vaan ensimm�isell� kerralla kun tauti iskee, siihen sopivaa informaatiota ei ole, vaan se on generoitava satunnaisin mutaatioin. Spetner on my�nt�nyt sen, ett� immuunisysteemi lis�� informaatiota B -solun genomiin. Kyse ei siis ole "potentiaali - informaation esiinkaivamisesta", vaan aidosta informaation kasvusta. Eli jotain, jonka piti h�nen omista m��ritelmist��n k�sin olla mahdotonta.
Spetner esitt�� seuraavia syit� sille,miksi t�m� on mahdollista:
*Mutaatiom��r� on suuri : Arviot v�lill� 100 miljoonaa - 10 miljardia kappaletta.
T�m�n p��telm�n Spetner perustelee, sivulla 103, sill� ett� todenn�k�isyys tuottaa informaatiota "evoluutiosattumalta" saadaan, kun kerrotaan jokaisen v�livaiheen todenn�k�isyydet kesken��n : T�m� todenn�k�isyys taas saavutetaan arvaamalla tarpeeksi monta kertaa. T�ss� n�kemyksess� kummallista on:
-Luonnonvalintaa ei oteta huomioon lainkaan, jolloin v�livaiheiden k�ytt� itse asiassa menett�� merkityksens�. V�livaiheella voi olla merkityst� vain luonnonvalinnan kautta. "V�livaiheilla" luodaan ainoastaan valheellinen mielikuva "evoluution huomioon ottamisessa".
-Miten t�t� selityst� soveltaen informaatiota ei synny luonnossa "miljoonissa vuosissa", kun todenn�k�isyydet pysyv�t ja mutaatiot tuottavat jatkuvasti "arvauksia"? Eik� sen pit�isi olla pikemminkin v�ist�m�t�nt�?
On joka tapauksessa eritt�in erikoista, ett� Spetner, jotka kiist�� "aidosti hy�dyllisten mutaatioiden" olemassaolonkin kaikesta huolimatta itse tarjoaa mutaatioita "tarpeeksi isoissa m��riss� esiintyess��n" selityksen� "aidosti uuden informaation" synnylle. Eih�n "puhtaan todenn�k�isyyden" t�yttymiselle, sellaisenahan Spetner tilanteen laskee- ole ratkaisevaa se, kuinka lyhyess� ajassa yritykset tapahtuvat, vaan siin� kuinka monta kertaa n�in tapahtuu.
*Immuunisysteemiss� k�sitelt�v� informaatiom��r� on pieni
T�m� on "virhekatastrofirajaan" sovellettuna aivan j�rkev� argumentti. Suuret mutaatiom��r�t kuitenkin toimisivat hajottavana voimana. Perim�ss�mme mutaatiom��r� on selv�sti immuunisysteemi� pienempi informaatiom��r� ovat selv�sti suurempi. N�in suurempi informaatiom��r� joudutaan kompensoimaan, eik� virhekatastrofiin ajauduta.
*Systeemi on hienos��detty.
On aivan totta, ett� informaatio "on biteiss�": sek� informaation kasvu ett� sen v�heneminen ovat muutoksia niiss� - luokiteltiin informaatio mill� tavoin tahansa. N�in ollen pelk�t muutokset eiv�t riit�, vaan tarvitaan jotain, joka "tarkastaa" mitk� ovat hyvi� ja mitk� huonoja muutoksia. Immuunisysteemiss� on tapoja tunnistaa hyv�t ja huonot muutokset, Spetner kutstuu t�t� "hienos��d�ksi". Luonnossa vastaavaa ei kutsuta "hienos��d�ksi" vaan "luonnonvalinnaksi" - ja t�t� evoluutikot pit�v�t kovasti arvossa.
*Muutokset eiv�t tallennu perim��n.
Spetnerin k�sitys muuttuu t�ll� perusteella hieman kummalliseksi, koska h�nen v�itteens� on ymm�rrett�viss� "Informaatiota ei voi synty�, jos lopputulos tallennetaan." Lis�ongelmia t�lle tuo se, ett� vaikka onkin totta, ett� immuunisysteemi ei tallenna muutoksia ituradan perim��n, niin silti tiedot tallentuvat immuunisysteemin soluihin: syntyy resistenssi ja sama tauti paranee seuraavalla kerralla nopeammin. Ilmeisesti pelkk� tallennuskaan ei siis voi olla este, vaan ilmeisesti jokin vaikutus on sill� minne se tallennus tehd��n.. Tai sitten t�m� argumentti on n�enn�inen.
Kun n�m� kaikki kohdat otetaan huomioon on helpohko huomata, ett� immuunisysteemi on "analoginen" evoluutioprosesseihin. Evoluutiokin on hienostunut systeemi, jossa ymp�rist� asettaa rajaukset informaatiolle, ja jossa esiintyy runsaasti mutaatioita ja jossa on valintamekanismi.
Yhteenvetoa
"The NREH, on the other hand, postulates nonrandom variation. It does not fulfill the neo-Darwinian agenda in that it does not contribute to a natural explanation of the origin of life."
(Lee Spetner, "Not by Chance!", 1998, sivu 210)
Spetnerin keskittyy evoluutiokritiikkiin, mutta muista poiketen esitt�� my�s vastahypoteesin, "NREH":n.
*Evoluutiohypoteesissaan Spetner nojaa omaan informaatiom��rittelyyns�, joka ei ole kovin osuva -Spetner ei esimerkiksi k�sit�, ett� entsyymien ja substraattien sitoutumisella ei ole "kaikki tai ei mit��n" -luonnetta. H�n my�s laittaa yht�l�isyysmerkit "puhtaan Sattuman" ja "evoluution" v�liin. Spetnerin k�ytt�m� "hienos��t�perustelu", joka sallii informaation kasvun taas on ominaisuuksiltaan kovasti samanlainen, kuin luonnonvalinta.
*NREH:ssa keskeisint� on "evoluution ennalta ohjelmoitu potentiaali", joka m��r�� miten kehitys menee ja mink�laista informaatiota tulee. Spetner ei kuitenkaan selit� miten mutaatiot kaivavat potentiaalista informaatiota, eik� sit� miten t�t� potentiaalia voitaisiin tutkia. H�n paljastaa, ett� osa on "nukkuvia geenej�", mutta t�m� ei ole ainut menetelm�. Spetner ei my�sk��n kerro, syntyyk� "NREH":n piiriss� uutta informaatiota vai ei. Emme loppupeleiss� tied� mik� t�m� "potentiaali" on, emmek� my�sk��n mitk� ovat "NREH":n rajat, ja t�t� kautta kautta meill� ei ole keinoa falsifioida Spetnerin teoriaa: Esimerkiksi havainnot, joissa Spetnerin m��ritelm�n mukainen informaatio lis��ntyy ei falsifioi h�nen malliaan, koska ne voidaan tulkita "tuntemattomasta potentiaalista" johtuviksi. Spetner ei my�sk��n sovella evoluutioon soveltamaansa "Voisiko-se-toimia?" -kaavaansa omaan informaatiomalliinsa: l�hinn� koska h�nen mallinsa ei ainakaan toistaiseksi kerro mit��n sellaista mit� evoluutiokaan ei kertoisi - h�nen mallinsa keskeisimm�t, eli "mutaatioita ohjaava voima" ja "mikroevoluution potentiaali" kun ovat molemmat paitsi tuntemattomia my�s tuntemattoman suuruisia.
Kaikesta huolimatta Spetnerin malli ansaitsee erikoishuomion siit�, ett� se on toistaiseksi ainut kreationistien informaatiomalli, joka on sek� formalisaatio ett� vaihtoehtoinen malli.
Lis�tietoja:
Timo Tiainen,"Kriittinen arvio Spetnerin n�kemyksist�",2004
Ian Musgrave,"Information Theory and Creationism - Spetner and Biological Information",2005
Gert Korthof,"Could it work?",2003
Mark Chu-Carroll, "Using Bad Math to Create Bad Models to Produce Bad Results",2007
Spetnerin kaavan mukaan informaatiom��r� kasvaa sit� mukaa, mit� enemm�n on olemassa substraatteja, joihin entsyymi ei vaikuta, joten uusia substraatteja synnytt�m�ll� informaatiom��r� voisi kasvaa: T�m� johtaa my�s siihen ett� sin� hetken� kun entsyymi menett�� kokonaan toimintansa, sill� ei ole substraattia ja entsyymin informaatio nousee "Spetnerin tasolla valtavaksi" koska substraattien m��r� joihin entsyymi ei vaikuta on "kaikki mahdolliset". T�lt� kannalta eli�, jonka sis�ll� ei olisi ainuttakaan toimivaa entsyymi� sis�lt�isi "maksimaalisesti informaatiota". -Joskin juuri t�m� ominaisuus selitt�� miten "nukkuva geeni" voi informaatiota kadottamalla muuttua toiminnalliseksi. Ennusteena Spetnerin teoriassa on, ett� satunnaiset mutaatiot tuhoaisivat informaatiota, jolloin satunnaismutaatiory�ppy aina kasvattaisi "entsyymien laaja -alaisuutta" - jolloin koetta jatkettaessa lopulta jokainen entsyymi toimisi jokaisen substraatin kanssa.
Spetnerin tunnetuimmat s��nn�t ovat "satunnaiset muutokset eiv�t tuota informaatiota" ja ett� "mutaatiot karsivat informaatiota"
[] => v [] a // satunnaisia muutoksia, m��r�isyys ei kasva.
a valikoituu // v + [] katoavat, informaatio v�henee
a => a ai ab // satunnaisia muutoksia, m��r�isyys ei kasva
ab => valikoituu // informaatio v�henee
...
a...y => a...z, a..ya //satunnaisia muutoksia, m��r�isyys ei kasva
a...z => valikoituu // informaatio v�henee
Spetnerlogiikalla informaation m��r� on koko ajan v�hentynyt, vaikka se on kasvanut nollasta 123:n bittiin ja informaatio on eksaktia. Salakuljetettua informaatiota siin� on samassa suhteessa miss� luonnossakin. Algoritmi tiet�� ainoastaan kuinka hyv� satunnaisten mutaatioiden muuntelema ratkaisu on tiettyjen kriteereiden valossa(mahdollisimman pitk� aakkosj�rjestys). Samoin luonnossa ymp�r�iv�t luonnonlait m��r��v�t sen mitk� muodot s�ilyv�t ja mitk� eiv�t.
Evoluution potentiaali
"A classic example is to remove the gene that allows E. coli bacteria to digest lactose, but then give the colony of bacteria only lactose to eat. Of the millions in a colony, some bacteria re-evolve the ability to hydrolyze lactose and survive."
(Science Daily Barry G. Hallin tutkimuksesta, "Darwin's Time Machine: Scientists Begin Predicting Evolution's Next Step", 20.Maal.2002.)
Informaationtuotto -ongelmien varalle Spetner on kehitt�nyt "mikroevoluution potentiaaliin" eli "esiohjelmoitu kehitys". T�ss� "eli�t ovat hienos��detyt", jolloin ilmestyy "yll�tt�v��kin muuntelua".
H�n vetoaa usein Barry G. Hallin 1982 tekem��n tutkimukseen. Siin� osoitettiin, ett� populaatioon ilmestyy s��nn�nmukaisesti ja sangen nopeasti tietyt muutokset tietyss� valintapaineessa. Spetnerin tulkinta kyseisest� tutkimuksesta on, ett� hy�dylliset mutaatiot ovat liian yleisi�, jolloin ne eiv�t ole "aidosti informaatiota luovia"(~Spetnerin "truly beneficial") vaan kaivavat informaation "potentiaalistaan".
Barry Hall on suorittanut sittemmin t�ydent�vi� tutkimuksia, mutta n�ist� Spetner ei puhu. Onhan se toisaalta ik�v��, ett� jo "klassisessa esimerkiss�" ensin suoritetaan "deleetio", eli koko ominaisuuden poisto ja se silti kehittyy takaisin - suorastaan ennustettavan pilkuntarkasti. "Potentiaali" ei siis voi olla perim�ss�. Vaikuttaa "evoluution hyvyysfunktioilta" eli tutummin "luonnonvalinnalta": T�ll�in Spetnerin "mikroevoluution potentiaali" tarkoittaa yksinkertaisesti samaa asiaa, mihin viittaa my�s Ernst Meyer 2003 julkaistun kirjansa "Evoluutio" sivulla 157 lauseessa "Koska kaikki kuviteltavissa olevat hy�dylliset mutaatiot ovat jo valikoituneet vakaissa elinymp�rist�iss�, uusien hy�dyllisten mutaatioiden syntyminen on melko harvinaista." Eli evoluutiolla on olemassa (lokaali)optimi, jonka "yli tai ohi" informaatiota ei voi synty�. K�yt�nn�ss� Spetnerin "neodarwinismin kritiikki" lakkaa olemasta "neodarwinismin kritiikki�", jos h�n my�nt�� luonnonvalinnan t�m�n ominaisuuden, joten ainut vaihtoehto on vetoaminen "nukkuviin geeneihin". Eli pelkk� klassinen "sokerijuurikkaan sokeripitoisuusesimerkki" tai muut vastaavat, jossa osoitetaan, ett� "informaatiolla on lokaali optimi" ei riit�: Ollakseen "evoluutiolle ongelma" t�ytyy my�s pysty� erottamaan "lokaali optimi" ja "mikroevoluution koko kyky" toisistaan ja osoittamaan, ett� kyse ei ole pelk�st� "lokaalista optimista": Lokaali optimi -v�ite kun on ongelmallinen siksi ett� vaikka evoluutioalgoritmeissa voi lokaali optimi olla pysyv�, luonnossa "hyvyysfunktiot" muuttuvat ajan kanssa. Jopa hyvin yksinkertainen esimerkki osoittaa sen ett� hyvyysfunktiot ja "lokaalit optimit" evoluutiossa ovat pakotettuja muuttumaan, ne eiv�t ole mik��n sein� kuin korkeintaan v�liaikaisesti:
Oletetaan nyt ett� meill� on hyvyysfunktio jossa on lokaali optimi, joka ei kuitenkaan ole "korkein mahdollinen huippu". Sitten l�yd�mme organismin, joka kiipe�� siihen. Kun evoluutio saavuttaa maksimin, evoluutio lakkaa etenem�st�, jonka seurauksena juuri sen lokaalin optimin kohdalla alkaa olemaan ruuhkaa: Yksil�t kuluttavat samaa ravintoa, joka taas vaikuttaa hyvyysfunktioon. Seurauksena lokaalin optimin paikka muuttuu.
Lis�ksi evoluutio on moniulotteinen, joten pit�isi todistaa ett� optimi olisi rajattu joka suuntaan. Jos meill� on hyvyysavaruus, jolla on X ja Y akseli, ei voida sanoa ett� se ett� kohde on sellaisella kohdalla ett� sen Y on lokaalissa optimissa, se ei tarkoita ett� se ei voisi muuttua paremmaksi, se voi liikkua X:n kautta, ja t�m� X:n muutos voi jopa tarjota uuden kohdan, jossa Y ei en�� olekaan sidottu. (Eli lukkiutumia voi synty� ja ne voivat vaihtaa paikkaa.) "Lokaali optimi" ei siksi toimi "mikroevoluutioesteen� jota ei voi ylitt��".
Eli "hyvyysfunktiot" ovat tuskin koskaan staattisia ja "useampi ulottuvuus" tuo lis�ksi uusia kiertoteit�.
Toisin sanoen, Spetnerist� s��nn�nmukainen kehitys ei suinkaan osoita, ett� h�nen omat laskelmansa "rappeutumisen pakosta" ja "evoluution satunnaisuudesta eli kyvytt�myydest� tuottaa ennusteita", olisivat v��r�ss�. H�n ei my�sk��n p���ttele, ett� "informaation trendi on nouseva", vaan sen sijaan se kertoo siit�, ett� muuntelumahdollisuuksien m��r� on rajallinen ja n�ist� vaihtoehdoista valikoituu tietty "sattumanvaraisesti eli ymp�rist�st� riippuen". Samanaikaisesti, kun "aktuaalinen, havaittava informaatio" kasvaa, niin "potentiaali pienenee".
T�lt� kohdin on eritt�in vaikeaa n�hd� eroa evoluutikkojen n�kemykseen ; heist� luonnonvalinta valikoi hyv�t ja huonot ominaisuudet ja ett� "jossain on potentiaalisessa muodossa tieto siit� millaista informaatiota eli�iss� voi olla". -Barry Hallin lis�ksi mm. Richard Dawkins on kirjoittanut luonnonvalinnan tuottamasta ennustettavuudesta- Evoluutikoista universumin sis�lt�m�t luonnonlait sek� n�iden keskin�inen vaikutus ymp�rist�ss� sis�lt�v�t "potentiaalisessa muodossa" tiedon "mink�lainen eli� on siihen sopiva ja mik� v�hemm�n sopiva". Spetner tulkitsee osan informaatiosta "nukkuviksi geeneiksi", mutta ei rajaa t�t� ainoaksi vaihtoehdoksi. Sill� miss� "potentiaali -informaation" varasto on, ei ole merkitt�v� asia. "Mikroevoluution potentiaali" voidaan siis varsin hyvin ymm�rt�� my�s "teistiseksi evoluutioksi" eik� oikein mitenk��n "evoluutiokritiikiksi".
Toisin sanoen Spetner ei vetoa ulkoisiin rakenteisiin. H�nen n�kemyksens� kannalta on t�ysin mahdollista, ett� koko eli�kunta olisi l�ht�isin esimerkiksi pienen bakteerin "potentiaalista". T�m� tarkoittaa sit�, ett� Spetner voi ajatella nykyisest� fossiilirekordista sen, ett� "potentiaali -informaatio" selv�sti ajan kanssa muuttuu "aktuaaliseksi informaatioksi".
T�ll� n�kemyksell� on muutamia heikkouksia:
*Ensinn�kin mink�lainen tahansa fossiiliaineisto sopii Spetnerin n�kemykseen, eli fossiileilla n�kemyst� ei voi falsifioida.
*Toisaalta t�m� johtaa mu�s siihen, ett� "rappeutuneiden rakenteiden" esittely ei my�sk��n ole mik��n todiste : Informaatiom��r� kun ei ole pelk�st��n n�kyviss� rakenteissa.
*Kolmanneksi jos potentiaali olisi koodattuna perim��n, ja se vain "nousisi sielt� ajan mittaan esiin", pit�isi mallin selitt�� miten; Ongelmana kun on se, ett� eitoiminnallinen, piilossa oleva, potentiaali on kuitenkin altis satunnaisille mutaatioille, jonka pit�isi johtaa siihen ett� ilman valintapainetta potentiaali tuhoutuu pitk�ll� t�ht�imell�: Toisin sanoen siin� miss� evoluutio EI v�it� ett� ominaisuudet nousisivat esiin sattumalta, vaan p�invastoin korostavat luonnonvalinnan merkityst� siin� ett� toiminta ei tuhoudu, koodattuun, eitoiminnalliseen, potentiaaliin uskova joutuisi hyv�ksym��n "sattumalta esiin nousevan informaation". Potentiaali voisi toki nousta esiin "eisatunnaisin mutaatioin", mutta t�ll�isest� manipulaatiosta ei ole havaintoja.
Spetnerin malli koskee pelk�st��n entsyymeit�, eik� siksi ole sovellettavissa rakenteisiin tai mihink��n muuhun. Spetnerin mallin falsifiointi on kyseenalaista, koska periaatteessa mik� tahansa informaatio voi olla per�isin "jostakin tuntemattomasta potentiaalista" ja vaikka jokin mutaatio muuttaisi jotain entsyymi� "spesifisemm�ksi", on se ainoastaan "spesifisemm�ksi suhteessa tiet�miimme substraatteihin", jolloin ei voida tiet�� onko kyse "aidosta spesifioitumisesta" eli "spesifisyydest� suhteessa kaikkiin substraatteihin". Jos potentiaali katsotaan rajoitetuksi pelk�st��n perim��n, niin tilanne muuttuu. Spetner muuttuu empiriaksi heti, kun tapa mitata potentiaalin suuruus keksit��n, ja heti, kun Spetner onnistuu keksim��n sen mekanismin, joka takaa sen, ett� mutaatiot ovatkin hy�dyllisi�, informaatiota "potentiaalista" esiinkaivavia, sen sijaan ett� olisivat "satunnaisia".
Mutaatiot - uuden rakentaminen vai varastostahaku
"I agree with him that these mutations add information to the B-cell genome."
(Lee Spetner, "Lee Spetner/Edward Max Dialogue", 2006)
On havaittu sellaisia "satunnaisia" mutaatioita, jotka lis��v�t geneettist� informaatiota jopa Spetnerin menetelm�ll� mitattuna. Erityisen vaikeaksi Spetnerin mallille on ollut immuunisysteemi, koska vasta -aineet eiv�t ole yhdistettyn� "nukkuviin geeneihin", vaan ensimm�isell� kerralla kun tauti iskee, siihen sopivaa informaatiota ei ole, vaan se on generoitava satunnaisin mutaatioin. Spetner on my�nt�nyt sen, ett� immuunisysteemi lis�� informaatiota B -solun genomiin. Kyse ei siis ole "potentiaali - informaation esiinkaivamisesta", vaan aidosta informaation kasvusta. Eli jotain, jonka piti h�nen omista m��ritelmist��n k�sin olla mahdotonta.
Spetner esitt�� seuraavia syit� sille,miksi t�m� on mahdollista:
*Mutaatiom��r� on suuri : Arviot v�lill� 100 miljoonaa - 10 miljardia kappaletta.
T�m�n p��telm�n Spetner perustelee, sivulla 103, sill� ett� todenn�k�isyys tuottaa informaatiota "evoluutiosattumalta" saadaan, kun kerrotaan jokaisen v�livaiheen todenn�k�isyydet kesken��n : T�m� todenn�k�isyys taas saavutetaan arvaamalla tarpeeksi monta kertaa. T�ss� n�kemyksess� kummallista on:
-Luonnonvalintaa ei oteta huomioon lainkaan, jolloin v�livaiheiden k�ytt� itse asiassa menett�� merkityksens�. V�livaiheella voi olla merkityst� vain luonnonvalinnan kautta. "V�livaiheilla" luodaan ainoastaan valheellinen mielikuva "evoluution huomioon ottamisessa".
-Miten t�t� selityst� soveltaen informaatiota ei synny luonnossa "miljoonissa vuosissa", kun todenn�k�isyydet pysyv�t ja mutaatiot tuottavat jatkuvasti "arvauksia"? Eik� sen pit�isi olla pikemminkin v�ist�m�t�nt�?
On joka tapauksessa eritt�in erikoista, ett� Spetner, jotka kiist�� "aidosti hy�dyllisten mutaatioiden" olemassaolonkin kaikesta huolimatta itse tarjoaa mutaatioita "tarpeeksi isoissa m��riss� esiintyess��n" selityksen� "aidosti uuden informaation" synnylle. Eih�n "puhtaan todenn�k�isyyden" t�yttymiselle, sellaisenahan Spetner tilanteen laskee- ole ratkaisevaa se, kuinka lyhyess� ajassa yritykset tapahtuvat, vaan siin� kuinka monta kertaa n�in tapahtuu.
*Immuunisysteemiss� k�sitelt�v� informaatiom��r� on pieni
T�m� on "virhekatastrofirajaan" sovellettuna aivan j�rkev� argumentti. Suuret mutaatiom��r�t kuitenkin toimisivat hajottavana voimana. Perim�ss�mme mutaatiom��r� on selv�sti immuunisysteemi� pienempi informaatiom��r� ovat selv�sti suurempi. N�in suurempi informaatiom��r� joudutaan kompensoimaan, eik� virhekatastrofiin ajauduta.
*Systeemi on hienos��detty.
On aivan totta, ett� informaatio "on biteiss�": sek� informaation kasvu ett� sen v�heneminen ovat muutoksia niiss� - luokiteltiin informaatio mill� tavoin tahansa. N�in ollen pelk�t muutokset eiv�t riit�, vaan tarvitaan jotain, joka "tarkastaa" mitk� ovat hyvi� ja mitk� huonoja muutoksia. Immuunisysteemiss� on tapoja tunnistaa hyv�t ja huonot muutokset, Spetner kutstuu t�t� "hienos��d�ksi". Luonnossa vastaavaa ei kutsuta "hienos��d�ksi" vaan "luonnonvalinnaksi" - ja t�t� evoluutikot pit�v�t kovasti arvossa.
*Muutokset eiv�t tallennu perim��n.
Spetnerin k�sitys muuttuu t�ll� perusteella hieman kummalliseksi, koska h�nen v�itteens� on ymm�rrett�viss� "Informaatiota ei voi synty�, jos lopputulos tallennetaan." Lis�ongelmia t�lle tuo se, ett� vaikka onkin totta, ett� immuunisysteemi ei tallenna muutoksia ituradan perim��n, niin silti tiedot tallentuvat immuunisysteemin soluihin: syntyy resistenssi ja sama tauti paranee seuraavalla kerralla nopeammin. Ilmeisesti pelkk� tallennuskaan ei siis voi olla este, vaan ilmeisesti jokin vaikutus on sill� minne se tallennus tehd��n.. Tai sitten t�m� argumentti on n�enn�inen.
Kun n�m� kaikki kohdat otetaan huomioon on helpohko huomata, ett� immuunisysteemi on "analoginen" evoluutioprosesseihin. Evoluutiokin on hienostunut systeemi, jossa ymp�rist� asettaa rajaukset informaatiolle, ja jossa esiintyy runsaasti mutaatioita ja jossa on valintamekanismi.
Yhteenvetoa
"The NREH, on the other hand, postulates nonrandom variation. It does not fulfill the neo-Darwinian agenda in that it does not contribute to a natural explanation of the origin of life."
(Lee Spetner, "Not by Chance!", 1998, sivu 210)
Spetnerin keskittyy evoluutiokritiikkiin, mutta muista poiketen esitt�� my�s vastahypoteesin, "NREH":n.
*Evoluutiohypoteesissaan Spetner nojaa omaan informaatiom��rittelyyns�, joka ei ole kovin osuva -Spetner ei esimerkiksi k�sit�, ett� entsyymien ja substraattien sitoutumisella ei ole "kaikki tai ei mit��n" -luonnetta. H�n my�s laittaa yht�l�isyysmerkit "puhtaan Sattuman" ja "evoluution" v�liin. Spetnerin k�ytt�m� "hienos��t�perustelu", joka sallii informaation kasvun taas on ominaisuuksiltaan kovasti samanlainen, kuin luonnonvalinta.
*NREH:ssa keskeisint� on "evoluution ennalta ohjelmoitu potentiaali", joka m��r�� miten kehitys menee ja mink�laista informaatiota tulee. Spetner ei kuitenkaan selit� miten mutaatiot kaivavat potentiaalista informaatiota, eik� sit� miten t�t� potentiaalia voitaisiin tutkia. H�n paljastaa, ett� osa on "nukkuvia geenej�", mutta t�m� ei ole ainut menetelm�. Spetner ei my�sk��n kerro, syntyyk� "NREH":n piiriss� uutta informaatiota vai ei. Emme loppupeleiss� tied� mik� t�m� "potentiaali" on, emmek� my�sk��n mitk� ovat "NREH":n rajat, ja t�t� kautta kautta meill� ei ole keinoa falsifioida Spetnerin teoriaa: Esimerkiksi havainnot, joissa Spetnerin m��ritelm�n mukainen informaatio lis��ntyy ei falsifioi h�nen malliaan, koska ne voidaan tulkita "tuntemattomasta potentiaalista" johtuviksi. Spetner ei my�sk��n sovella evoluutioon soveltamaansa "Voisiko-se-toimia?" -kaavaansa omaan informaatiomalliinsa: l�hinn� koska h�nen mallinsa ei ainakaan toistaiseksi kerro mit��n sellaista mit� evoluutiokaan ei kertoisi - h�nen mallinsa keskeisimm�t, eli "mutaatioita ohjaava voima" ja "mikroevoluution potentiaali" kun ovat molemmat paitsi tuntemattomia my�s tuntemattoman suuruisia.
Kaikesta huolimatta Spetnerin malli ansaitsee erikoishuomion siit�, ett� se on toistaiseksi ainut kreationistien informaatiomalli, joka on sek� formalisaatio ett� vaihtoehtoinen malli.
Lis�tietoja:
Timo Tiainen,"Kriittinen arvio Spetnerin n�kemyksist�",2004
Ian Musgrave,"Information Theory and Creationism - Spetner and Biological Information",2005
Gert Korthof,"Could it work?",2003
Mark Chu-Carroll, "Using Bad Math to Create Bad Models to Produce Bad Results",2007
Evoluution potentiaali
"A classic example is to remove the gene that allows E. coli bacteria to digest lactose, but then give the colony of bacteria only lactose to eat. Of the millions in a colony, some bacteria re-evolve the ability to hydrolyze lactose and survive."
(Science Daily Barry G. Hallin tutkimuksesta, "Darwin's Time Machine: Scientists Begin Predicting Evolution's Next Step", 20.Maal.2002.)
Informaationtuotto -ongelmien varalle Spetner on kehitt�nyt "mikroevoluution potentiaaliin" eli "esiohjelmoitu kehitys". T�ss� "eli�t ovat hienos��detyt", jolloin ilmestyy "yll�tt�v��kin muuntelua".
H�n vetoaa usein Barry G. Hallin 1982 tekem��n tutkimukseen. Siin� osoitettiin, ett� populaatioon ilmestyy s��nn�nmukaisesti ja sangen nopeasti tietyt muutokset tietyss� valintapaineessa. Spetnerin tulkinta kyseisest� tutkimuksesta on, ett� hy�dylliset mutaatiot ovat liian yleisi�, jolloin ne eiv�t ole "aidosti informaatiota luovia"(~Spetnerin "truly beneficial") vaan kaivavat informaation "potentiaalistaan".
Barry Hall on suorittanut sittemmin t�ydent�vi� tutkimuksia, mutta n�ist� Spetner ei puhu. Onhan se toisaalta ik�v��, ett� jo "klassisessa esimerkiss�" ensin suoritetaan "deleetio", eli koko ominaisuuden poisto ja se silti kehittyy takaisin - suorastaan ennustettavan pilkuntarkasti. "Potentiaali" ei siis voi olla perim�ss�. Vaikuttaa "evoluution hyvyysfunktioilta" eli tutummin "luonnonvalinnalta": T�ll�in Spetnerin "mikroevoluution potentiaali" tarkoittaa yksinkertaisesti samaa asiaa, mihin viittaa my�s Ernst Meyer 2003 julkaistun kirjansa "Evoluutio" sivulla 157 lauseessa "Koska kaikki kuviteltavissa olevat hy�dylliset mutaatiot ovat jo valikoituneet vakaissa elinymp�rist�iss�, uusien hy�dyllisten mutaatioiden syntyminen on melko harvinaista." Eli evoluutiolla on olemassa (lokaali)optimi, jonka "yli tai ohi" informaatiota ei voi synty�. K�yt�nn�ss� Spetnerin "neodarwinismin kritiikki" lakkaa olemasta "neodarwinismin kritiikki�", jos h�n my�nt�� luonnonvalinnan t�m�n ominaisuuden, joten ainut vaihtoehto on vetoaminen "nukkuviin geeneihin". Eli pelkk� klassinen "sokerijuurikkaan sokeripitoisuusesimerkki" tai muut vastaavat, jossa osoitetaan, ett� "informaatiolla on lokaali optimi" ei riit�: Ollakseen "evoluutiolle ongelma" t�ytyy my�s pysty� erottamaan "lokaali optimi" ja "mikroevoluution koko kyky" toisistaan ja osoittamaan, ett� kyse ei ole pelk�st� "lokaalista optimista": Lokaali optimi -v�ite kun on ongelmallinen siksi ett� vaikka evoluutioalgoritmeissa voi lokaali optimi olla pysyv�, luonnossa "hyvyysfunktiot" muuttuvat ajan kanssa. Jopa hyvin yksinkertainen esimerkki osoittaa sen ett� hyvyysfunktiot ja "lokaalit optimit" evoluutiossa ovat pakotettuja muuttumaan, ne eiv�t ole mik��n sein� kuin korkeintaan v�liaikaisesti:
Oletetaan nyt ett� meill� on hyvyysfunktio jossa on lokaali optimi, joka ei kuitenkaan ole "korkein mahdollinen huippu". Sitten l�yd�mme organismin, joka kiipe�� siihen. Kun evoluutio saavuttaa maksimin, evoluutio lakkaa etenem�st�, jonka seurauksena juuri sen lokaalin optimin kohdalla alkaa olemaan ruuhkaa: Yksil�t kuluttavat samaa ravintoa, joka taas vaikuttaa hyvyysfunktioon. Seurauksena lokaalin optimin paikka muuttuu.
Lis�ksi evoluutio on moniulotteinen, joten pit�isi todistaa ett� optimi olisi rajattu joka suuntaan. Jos meill� on hyvyysavaruus, jolla on X ja Y akseli, ei voida sanoa ett� se ett� kohde on sellaisella kohdalla ett� sen Y on lokaalissa optimissa, se ei tarkoita ett� se ei voisi muuttua paremmaksi, se voi liikkua X:n kautta, ja t�m� X:n muutos voi jopa tarjota uuden kohdan, jossa Y ei en�� olekaan sidottu. (Eli lukkiutumia voi synty� ja ne voivat vaihtaa paikkaa.) "Lokaali optimi" ei siksi toimi "mikroevoluutioesteen� jota ei voi ylitt��".
Eli "hyvyysfunktiot" ovat tuskin koskaan staattisia ja "useampi ulottuvuus" tuo lis�ksi uusia kiertoteit�.
Toisin sanoen, Spetnerist� s��nn�nmukainen kehitys ei suinkaan osoita, ett� h�nen omat laskelmansa "rappeutumisen pakosta" ja "evoluution satunnaisuudesta eli kyvytt�myydest� tuottaa ennusteita", olisivat v��r�ss�. H�n ei my�sk��n p���ttele, ett� "informaation trendi on nouseva", vaan sen sijaan se kertoo siit�, ett� muuntelumahdollisuuksien m��r� on rajallinen ja n�ist� vaihtoehdoista valikoituu tietty "sattumanvaraisesti eli ymp�rist�st� riippuen". Samanaikaisesti, kun "aktuaalinen, havaittava informaatio" kasvaa, niin "potentiaali pienenee".
T�lt� kohdin on eritt�in vaikeaa n�hd� eroa evoluutikkojen n�kemykseen ; heist� luonnonvalinta valikoi hyv�t ja huonot ominaisuudet ja ett� "jossain on potentiaalisessa muodossa tieto siit� millaista informaatiota eli�iss� voi olla". -Barry Hallin lis�ksi mm. Richard Dawkins on kirjoittanut luonnonvalinnan tuottamasta ennustettavuudesta- Evoluutikoista universumin sis�lt�m�t luonnonlait sek� n�iden keskin�inen vaikutus ymp�rist�ss� sis�lt�v�t "potentiaalisessa muodossa" tiedon "mink�lainen eli� on siihen sopiva ja mik� v�hemm�n sopiva". Spetner tulkitsee osan informaatiosta "nukkuviksi geeneiksi", mutta ei rajaa t�t� ainoaksi vaihtoehdoksi. Sill� miss� "potentiaali -informaation" varasto on, ei ole merkitt�v� asia. "Mikroevoluution potentiaali" voidaan siis varsin hyvin ymm�rt�� my�s "teistiseksi evoluutioksi" eik� oikein mitenk��n "evoluutiokritiikiksi".
Toisin sanoen Spetner ei vetoa ulkoisiin rakenteisiin. H�nen n�kemyksens� kannalta on t�ysin mahdollista, ett� koko eli�kunta olisi l�ht�isin esimerkiksi pienen bakteerin "potentiaalista". T�m� tarkoittaa sit�, ett� Spetner voi ajatella nykyisest� fossiilirekordista sen, ett� "potentiaali -informaatio" selv�sti ajan kanssa muuttuu "aktuaaliseksi informaatioksi".
T�ll� n�kemyksell� on muutamia heikkouksia:
*Ensinn�kin mink�lainen tahansa fossiiliaineisto sopii Spetnerin n�kemykseen, eli fossiileilla n�kemyst� ei voi falsifioida.
*Toisaalta t�m� johtaa mu�s siihen, ett� "rappeutuneiden rakenteiden" esittely ei my�sk��n ole mik��n todiste : Informaatiom��r� kun ei ole pelk�st��n n�kyviss� rakenteissa.
*Kolmanneksi jos potentiaali olisi koodattuna perim��n, ja se vain "nousisi sielt� ajan mittaan esiin", pit�isi mallin selitt�� miten; Ongelmana kun on se, ett� eitoiminnallinen, piilossa oleva, potentiaali on kuitenkin altis satunnaisille mutaatioille, jonka pit�isi johtaa siihen ett� ilman valintapainetta potentiaali tuhoutuu pitk�ll� t�ht�imell�: Toisin sanoen siin� miss� evoluutio EI v�it� ett� ominaisuudet nousisivat esiin sattumalta, vaan p�invastoin korostavat luonnonvalinnan merkityst� siin� ett� toiminta ei tuhoudu, koodattuun, eitoiminnalliseen, potentiaaliin uskova joutuisi hyv�ksym��n "sattumalta esiin nousevan informaation". Potentiaali voisi toki nousta esiin "eisatunnaisin mutaatioin", mutta t�ll�isest� manipulaatiosta ei ole havaintoja.
Spetnerin malli koskee pelk�st��n entsyymeit�, eik� siksi ole sovellettavissa rakenteisiin tai mihink��n muuhun. Spetnerin mallin falsifiointi on kyseenalaista, koska periaatteessa mik� tahansa informaatio voi olla per�isin "jostakin tuntemattomasta potentiaalista" ja vaikka jokin mutaatio muuttaisi jotain entsyymi� "spesifisemm�ksi", on se ainoastaan "spesifisemm�ksi suhteessa tiet�miimme substraatteihin", jolloin ei voida tiet�� onko kyse "aidosta spesifioitumisesta" eli "spesifisyydest� suhteessa kaikkiin substraatteihin". Jos potentiaali katsotaan rajoitetuksi pelk�st��n perim��n, niin tilanne muuttuu. Spetner muuttuu empiriaksi heti, kun tapa mitata potentiaalin suuruus keksit��n, ja heti, kun Spetner onnistuu keksim��n sen mekanismin, joka takaa sen, ett� mutaatiot ovatkin hy�dyllisi�, informaatiota "potentiaalista" esiinkaivavia, sen sijaan ett� olisivat "satunnaisia".
Mutaatiot - uuden rakentaminen vai varastostahaku
"I agree with him that these mutations add information to the B-cell genome."
(Lee Spetner, "Lee Spetner/Edward Max Dialogue", 2006)
On havaittu sellaisia "satunnaisia" mutaatioita, jotka lis��v�t geneettist� informaatiota jopa Spetnerin menetelm�ll� mitattuna. Erityisen vaikeaksi Spetnerin mallille on ollut immuunisysteemi, koska vasta -aineet eiv�t ole yhdistettyn� "nukkuviin geeneihin", vaan ensimm�isell� kerralla kun tauti iskee, siihen sopivaa informaatiota ei ole, vaan se on generoitava satunnaisin mutaatioin. Spetner on my�nt�nyt sen, ett� immuunisysteemi lis�� informaatiota B -solun genomiin. Kyse ei siis ole "potentiaali - informaation esiinkaivamisesta", vaan aidosta informaation kasvusta. Eli jotain, jonka piti h�nen omista m��ritelmist��n k�sin olla mahdotonta.
Spetner esitt�� seuraavia syit� sille,miksi t�m� on mahdollista:
*Mutaatiom��r� on suuri : Arviot v�lill� 100 miljoonaa - 10 miljardia kappaletta.
T�m�n p��telm�n Spetner perustelee, sivulla 103, sill� ett� todenn�k�isyys tuottaa informaatiota "evoluutiosattumalta" saadaan, kun kerrotaan jokaisen v�livaiheen todenn�k�isyydet kesken��n : T�m� todenn�k�isyys taas saavutetaan arvaamalla tarpeeksi monta kertaa. T�ss� n�kemyksess� kummallista on:
-Luonnonvalintaa ei oteta huomioon lainkaan, jolloin v�livaiheiden k�ytt� itse asiassa menett�� merkityksens�. V�livaiheella voi olla merkityst� vain luonnonvalinnan kautta. "V�livaiheilla" luodaan ainoastaan valheellinen mielikuva "evoluution huomioon ottamisessa".
-Miten t�t� selityst� soveltaen informaatiota ei synny luonnossa "miljoonissa vuosissa", kun todenn�k�isyydet pysyv�t ja mutaatiot tuottavat jatkuvasti "arvauksia"? Eik� sen pit�isi olla pikemminkin v�ist�m�t�nt�?
On joka tapauksessa eritt�in erikoista, ett� Spetner, jotka kiist�� "aidosti hy�dyllisten mutaatioiden" olemassaolonkin kaikesta huolimatta itse tarjoaa mutaatioita "tarpeeksi isoissa m��riss� esiintyess��n" selityksen� "aidosti uuden informaation" synnylle. Eih�n "puhtaan todenn�k�isyyden" t�yttymiselle, sellaisenahan Spetner tilanteen laskee- ole ratkaisevaa se, kuinka lyhyess� ajassa yritykset tapahtuvat, vaan siin� kuinka monta kertaa n�in tapahtuu.
*Immuunisysteemiss� k�sitelt�v� informaatiom��r� on pieni
T�m� on "virhekatastrofirajaan" sovellettuna aivan j�rkev� argumentti. Suuret mutaatiom��r�t kuitenkin toimisivat hajottavana voimana. Perim�ss�mme mutaatiom��r� on selv�sti immuunisysteemi� pienempi informaatiom��r� ovat selv�sti suurempi. N�in suurempi informaatiom��r� joudutaan kompensoimaan, eik� virhekatastrofiin ajauduta.
*Systeemi on hienos��detty.
On aivan totta, ett� informaatio "on biteiss�": sek� informaation kasvu ett� sen v�heneminen ovat muutoksia niiss� - luokiteltiin informaatio mill� tavoin tahansa. N�in ollen pelk�t muutokset eiv�t riit�, vaan tarvitaan jotain, joka "tarkastaa" mitk� ovat hyvi� ja mitk� huonoja muutoksia. Immuunisysteemiss� on tapoja tunnistaa hyv�t ja huonot muutokset, Spetner kutstuu t�t� "hienos��d�ksi". Luonnossa vastaavaa ei kutsuta "hienos��d�ksi" vaan "luonnonvalinnaksi" - ja t�t� evoluutikot pit�v�t kovasti arvossa.
*Muutokset eiv�t tallennu perim��n.
Spetnerin k�sitys muuttuu t�ll� perusteella hieman kummalliseksi, koska h�nen v�itteens� on ymm�rrett�viss� "Informaatiota ei voi synty�, jos lopputulos tallennetaan." Lis�ongelmia t�lle tuo se, ett� vaikka onkin totta, ett� immuunisysteemi ei tallenna muutoksia ituradan perim��n, niin silti tiedot tallentuvat immuunisysteemin soluihin: syntyy resistenssi ja sama tauti paranee seuraavalla kerralla nopeammin. Ilmeisesti pelkk� tallennuskaan ei siis voi olla este, vaan ilmeisesti jokin vaikutus on sill� minne se tallennus tehd��n.. Tai sitten t�m� argumentti on n�enn�inen.
Kun n�m� kaikki kohdat otetaan huomioon on helpohko huomata, ett� immuunisysteemi on "analoginen" evoluutioprosesseihin. Evoluutiokin on hienostunut systeemi, jossa ymp�rist� asettaa rajaukset informaatiolle, ja jossa esiintyy runsaasti mutaatioita ja jossa on valintamekanismi.
Yhteenvetoa
"The NREH, on the other hand, postulates nonrandom variation. It does not fulfill the neo-Darwinian agenda in that it does not contribute to a natural explanation of the origin of life."
(Lee Spetner, "Not by Chance!", 1998, sivu 210)
Spetnerin keskittyy evoluutiokritiikkiin, mutta muista poiketen esitt�� my�s vastahypoteesin, "NREH":n.
*Evoluutiohypoteesissaan Spetner nojaa omaan informaatiom��rittelyyns�, joka ei ole kovin osuva -Spetner ei esimerkiksi k�sit�, ett� entsyymien ja substraattien sitoutumisella ei ole "kaikki tai ei mit��n" -luonnetta. H�n my�s laittaa yht�l�isyysmerkit "puhtaan Sattuman" ja "evoluution" v�liin. Spetnerin k�ytt�m� "hienos��t�perustelu", joka sallii informaation kasvun taas on ominaisuuksiltaan kovasti samanlainen, kuin luonnonvalinta.
*NREH:ssa keskeisint� on "evoluution ennalta ohjelmoitu potentiaali", joka m��r�� miten kehitys menee ja mink�laista informaatiota tulee. Spetner ei kuitenkaan selit� miten mutaatiot kaivavat potentiaalista informaatiota, eik� sit� miten t�t� potentiaalia voitaisiin tutkia. H�n paljastaa, ett� osa on "nukkuvia geenej�", mutta t�m� ei ole ainut menetelm�. Spetner ei my�sk��n kerro, syntyyk� "NREH":n piiriss� uutta informaatiota vai ei. Emme loppupeleiss� tied� mik� t�m� "potentiaali" on, emmek� my�sk��n mitk� ovat "NREH":n rajat, ja t�t� kautta kautta meill� ei ole keinoa falsifioida Spetnerin teoriaa: Esimerkiksi havainnot, joissa Spetnerin m��ritelm�n mukainen informaatio lis��ntyy ei falsifioi h�nen malliaan, koska ne voidaan tulkita "tuntemattomasta potentiaalista" johtuviksi. Spetner ei my�sk��n sovella evoluutioon soveltamaansa "Voisiko-se-toimia?" -kaavaansa omaan informaatiomalliinsa: l�hinn� koska h�nen mallinsa ei ainakaan toistaiseksi kerro mit��n sellaista mit� evoluutiokaan ei kertoisi - h�nen mallinsa keskeisimm�t, eli "mutaatioita ohjaava voima" ja "mikroevoluution potentiaali" kun ovat molemmat paitsi tuntemattomia my�s tuntemattoman suuruisia.
Kaikesta huolimatta Spetnerin malli ansaitsee erikoishuomion siit�, ett� se on toistaiseksi ainut kreationistien informaatiomalli, joka on sek� formalisaatio ett� vaihtoehtoinen malli.
Lis�tietoja:
Timo Tiainen,"Kriittinen arvio Spetnerin n�kemyksist�",2004
Ian Musgrave,"Information Theory and Creationism - Spetner and Biological Information",2005
Gert Korthof,"Could it work?",2003
Mark Chu-Carroll, "Using Bad Math to Create Bad Models to Produce Bad Results",2007
(Science Daily Barry G. Hallin tutkimuksesta, "Darwin's Time Machine: Scientists Begin Predicting Evolution's Next Step", 20.Maal.2002.)
Informaationtuotto -ongelmien varalle Spetner on kehitt�nyt "mikroevoluution potentiaaliin" eli "esiohjelmoitu kehitys". T�ss� "eli�t ovat hienos��detyt", jolloin ilmestyy "yll�tt�v��kin muuntelua".
H�n vetoaa usein Barry G. Hallin 1982 tekem��n tutkimukseen. Siin� osoitettiin, ett� populaatioon ilmestyy s��nn�nmukaisesti ja sangen nopeasti tietyt muutokset tietyss� valintapaineessa. Spetnerin tulkinta kyseisest� tutkimuksesta on, ett� hy�dylliset mutaatiot ovat liian yleisi�, jolloin ne eiv�t ole "aidosti informaatiota luovia"(~Spetnerin "truly beneficial") vaan kaivavat informaation "potentiaalistaan".
Barry Hall on suorittanut sittemmin t�ydent�vi� tutkimuksia, mutta n�ist� Spetner ei puhu. Onhan se toisaalta ik�v��, ett� jo "klassisessa esimerkiss�" ensin suoritetaan "deleetio", eli koko ominaisuuden poisto ja se silti kehittyy takaisin - suorastaan ennustettavan pilkuntarkasti. "Potentiaali" ei siis voi olla perim�ss�. Vaikuttaa "evoluution hyvyysfunktioilta" eli tutummin "luonnonvalinnalta": T�ll�in Spetnerin "mikroevoluution potentiaali" tarkoittaa yksinkertaisesti samaa asiaa, mihin viittaa my�s Ernst Meyer 2003 julkaistun kirjansa "Evoluutio" sivulla 157 lauseessa "Koska kaikki kuviteltavissa olevat hy�dylliset mutaatiot ovat jo valikoituneet vakaissa elinymp�rist�iss�, uusien hy�dyllisten mutaatioiden syntyminen on melko harvinaista." Eli evoluutiolla on olemassa (lokaali)optimi, jonka "yli tai ohi" informaatiota ei voi synty�. K�yt�nn�ss� Spetnerin "neodarwinismin kritiikki" lakkaa olemasta "neodarwinismin kritiikki�", jos h�n my�nt�� luonnonvalinnan t�m�n ominaisuuden, joten ainut vaihtoehto on vetoaminen "nukkuviin geeneihin". Eli pelkk� klassinen "sokerijuurikkaan sokeripitoisuusesimerkki" tai muut vastaavat, jossa osoitetaan, ett� "informaatiolla on lokaali optimi" ei riit�: Ollakseen "evoluutiolle ongelma" t�ytyy my�s pysty� erottamaan "lokaali optimi" ja "mikroevoluution koko kyky" toisistaan ja osoittamaan, ett� kyse ei ole pelk�st� "lokaalista optimista": Lokaali optimi -v�ite kun on ongelmallinen siksi ett� vaikka evoluutioalgoritmeissa voi lokaali optimi olla pysyv�, luonnossa "hyvyysfunktiot" muuttuvat ajan kanssa. Jopa hyvin yksinkertainen esimerkki osoittaa sen ett� hyvyysfunktiot ja "lokaalit optimit" evoluutiossa ovat pakotettuja muuttumaan, ne eiv�t ole mik��n sein� kuin korkeintaan v�liaikaisesti:
Oletetaan nyt ett� meill� on hyvyysfunktio jossa on lokaali optimi, joka ei kuitenkaan ole "korkein mahdollinen huippu". Sitten l�yd�mme organismin, joka kiipe�� siihen. Kun evoluutio saavuttaa maksimin, evoluutio lakkaa etenem�st�, jonka seurauksena juuri sen lokaalin optimin kohdalla alkaa olemaan ruuhkaa: Yksil�t kuluttavat samaa ravintoa, joka taas vaikuttaa hyvyysfunktioon. Seurauksena lokaalin optimin paikka muuttuu.
Lis�ksi evoluutio on moniulotteinen, joten pit�isi todistaa ett� optimi olisi rajattu joka suuntaan. Jos meill� on hyvyysavaruus, jolla on X ja Y akseli, ei voida sanoa ett� se ett� kohde on sellaisella kohdalla ett� sen Y on lokaalissa optimissa, se ei tarkoita ett� se ei voisi muuttua paremmaksi, se voi liikkua X:n kautta, ja t�m� X:n muutos voi jopa tarjota uuden kohdan, jossa Y ei en�� olekaan sidottu. (Eli lukkiutumia voi synty� ja ne voivat vaihtaa paikkaa.) "Lokaali optimi" ei siksi toimi "mikroevoluutioesteen� jota ei voi ylitt��".
Eli "hyvyysfunktiot" ovat tuskin koskaan staattisia ja "useampi ulottuvuus" tuo lis�ksi uusia kiertoteit�.
Toisin sanoen, Spetnerist� s��nn�nmukainen kehitys ei suinkaan osoita, ett� h�nen omat laskelmansa "rappeutumisen pakosta" ja "evoluution satunnaisuudesta eli kyvytt�myydest� tuottaa ennusteita", olisivat v��r�ss�. H�n ei my�sk��n p���ttele, ett� "informaation trendi on nouseva", vaan sen sijaan se kertoo siit�, ett� muuntelumahdollisuuksien m��r� on rajallinen ja n�ist� vaihtoehdoista valikoituu tietty "sattumanvaraisesti eli ymp�rist�st� riippuen". Samanaikaisesti, kun "aktuaalinen, havaittava informaatio" kasvaa, niin "potentiaali pienenee".
T�lt� kohdin on eritt�in vaikeaa n�hd� eroa evoluutikkojen n�kemykseen ; heist� luonnonvalinta valikoi hyv�t ja huonot ominaisuudet ja ett� "jossain on potentiaalisessa muodossa tieto siit� millaista informaatiota eli�iss� voi olla". -Barry Hallin lis�ksi mm. Richard Dawkins on kirjoittanut luonnonvalinnan tuottamasta ennustettavuudesta- Evoluutikoista universumin sis�lt�m�t luonnonlait sek� n�iden keskin�inen vaikutus ymp�rist�ss� sis�lt�v�t "potentiaalisessa muodossa" tiedon "mink�lainen eli� on siihen sopiva ja mik� v�hemm�n sopiva". Spetner tulkitsee osan informaatiosta "nukkuviksi geeneiksi", mutta ei rajaa t�t� ainoaksi vaihtoehdoksi. Sill� miss� "potentiaali -informaation" varasto on, ei ole merkitt�v� asia. "Mikroevoluution potentiaali" voidaan siis varsin hyvin ymm�rt�� my�s "teistiseksi evoluutioksi" eik� oikein mitenk��n "evoluutiokritiikiksi".
Toisin sanoen Spetner ei vetoa ulkoisiin rakenteisiin. H�nen n�kemyksens� kannalta on t�ysin mahdollista, ett� koko eli�kunta olisi l�ht�isin esimerkiksi pienen bakteerin "potentiaalista". T�m� tarkoittaa sit�, ett� Spetner voi ajatella nykyisest� fossiilirekordista sen, ett� "potentiaali -informaatio" selv�sti ajan kanssa muuttuu "aktuaaliseksi informaatioksi".
T�ll� n�kemyksell� on muutamia heikkouksia:
*Ensinn�kin mink�lainen tahansa fossiiliaineisto sopii Spetnerin n�kemykseen, eli fossiileilla n�kemyst� ei voi falsifioida.
*Toisaalta t�m� johtaa mu�s siihen, ett� "rappeutuneiden rakenteiden" esittely ei my�sk��n ole mik��n todiste : Informaatiom��r� kun ei ole pelk�st��n n�kyviss� rakenteissa.
*Kolmanneksi jos potentiaali olisi koodattuna perim��n, ja se vain "nousisi sielt� ajan mittaan esiin", pit�isi mallin selitt�� miten; Ongelmana kun on se, ett� eitoiminnallinen, piilossa oleva, potentiaali on kuitenkin altis satunnaisille mutaatioille, jonka pit�isi johtaa siihen ett� ilman valintapainetta potentiaali tuhoutuu pitk�ll� t�ht�imell�: Toisin sanoen siin� miss� evoluutio EI v�it� ett� ominaisuudet nousisivat esiin sattumalta, vaan p�invastoin korostavat luonnonvalinnan merkityst� siin� ett� toiminta ei tuhoudu, koodattuun, eitoiminnalliseen, potentiaaliin uskova joutuisi hyv�ksym��n "sattumalta esiin nousevan informaation". Potentiaali voisi toki nousta esiin "eisatunnaisin mutaatioin", mutta t�ll�isest� manipulaatiosta ei ole havaintoja.
Spetnerin malli koskee pelk�st��n entsyymeit�, eik� siksi ole sovellettavissa rakenteisiin tai mihink��n muuhun. Spetnerin mallin falsifiointi on kyseenalaista, koska periaatteessa mik� tahansa informaatio voi olla per�isin "jostakin tuntemattomasta potentiaalista" ja vaikka jokin mutaatio muuttaisi jotain entsyymi� "spesifisemm�ksi", on se ainoastaan "spesifisemm�ksi suhteessa tiet�miimme substraatteihin", jolloin ei voida tiet�� onko kyse "aidosta spesifioitumisesta" eli "spesifisyydest� suhteessa kaikkiin substraatteihin". Jos potentiaali katsotaan rajoitetuksi pelk�st��n perim��n, niin tilanne muuttuu. Spetner muuttuu empiriaksi heti, kun tapa mitata potentiaalin suuruus keksit��n, ja heti, kun Spetner onnistuu keksim��n sen mekanismin, joka takaa sen, ett� mutaatiot ovatkin hy�dyllisi�, informaatiota "potentiaalista" esiinkaivavia, sen sijaan ett� olisivat "satunnaisia".
Mutaatiot - uuden rakentaminen vai varastostahaku
"I agree with him that these mutations add information to the B-cell genome."
(Lee Spetner, "Lee Spetner/Edward Max Dialogue", 2006)
On havaittu sellaisia "satunnaisia" mutaatioita, jotka lis��v�t geneettist� informaatiota jopa Spetnerin menetelm�ll� mitattuna. Erityisen vaikeaksi Spetnerin mallille on ollut immuunisysteemi, koska vasta -aineet eiv�t ole yhdistettyn� "nukkuviin geeneihin", vaan ensimm�isell� kerralla kun tauti iskee, siihen sopivaa informaatiota ei ole, vaan se on generoitava satunnaisin mutaatioin. Spetner on my�nt�nyt sen, ett� immuunisysteemi lis�� informaatiota B -solun genomiin. Kyse ei siis ole "potentiaali - informaation esiinkaivamisesta", vaan aidosta informaation kasvusta. Eli jotain, jonka piti h�nen omista m��ritelmist��n k�sin olla mahdotonta.
Spetner esitt�� seuraavia syit� sille,miksi t�m� on mahdollista:
*Mutaatiom��r� on suuri : Arviot v�lill� 100 miljoonaa - 10 miljardia kappaletta.
T�m�n p��telm�n Spetner perustelee, sivulla 103, sill� ett� todenn�k�isyys tuottaa informaatiota "evoluutiosattumalta" saadaan, kun kerrotaan jokaisen v�livaiheen todenn�k�isyydet kesken��n : T�m� todenn�k�isyys taas saavutetaan arvaamalla tarpeeksi monta kertaa. T�ss� n�kemyksess� kummallista on:
-Luonnonvalintaa ei oteta huomioon lainkaan, jolloin v�livaiheiden k�ytt� itse asiassa menett�� merkityksens�. V�livaiheella voi olla merkityst� vain luonnonvalinnan kautta. "V�livaiheilla" luodaan ainoastaan valheellinen mielikuva "evoluution huomioon ottamisessa".
-Miten t�t� selityst� soveltaen informaatiota ei synny luonnossa "miljoonissa vuosissa", kun todenn�k�isyydet pysyv�t ja mutaatiot tuottavat jatkuvasti "arvauksia"? Eik� sen pit�isi olla pikemminkin v�ist�m�t�nt�?
On joka tapauksessa eritt�in erikoista, ett� Spetner, jotka kiist�� "aidosti hy�dyllisten mutaatioiden" olemassaolonkin kaikesta huolimatta itse tarjoaa mutaatioita "tarpeeksi isoissa m��riss� esiintyess��n" selityksen� "aidosti uuden informaation" synnylle. Eih�n "puhtaan todenn�k�isyyden" t�yttymiselle, sellaisenahan Spetner tilanteen laskee- ole ratkaisevaa se, kuinka lyhyess� ajassa yritykset tapahtuvat, vaan siin� kuinka monta kertaa n�in tapahtuu.
*Immuunisysteemiss� k�sitelt�v� informaatiom��r� on pieni
T�m� on "virhekatastrofirajaan" sovellettuna aivan j�rkev� argumentti. Suuret mutaatiom��r�t kuitenkin toimisivat hajottavana voimana. Perim�ss�mme mutaatiom��r� on selv�sti immuunisysteemi� pienempi informaatiom��r� ovat selv�sti suurempi. N�in suurempi informaatiom��r� joudutaan kompensoimaan, eik� virhekatastrofiin ajauduta.
*Systeemi on hienos��detty.
On aivan totta, ett� informaatio "on biteiss�": sek� informaation kasvu ett� sen v�heneminen ovat muutoksia niiss� - luokiteltiin informaatio mill� tavoin tahansa. N�in ollen pelk�t muutokset eiv�t riit�, vaan tarvitaan jotain, joka "tarkastaa" mitk� ovat hyvi� ja mitk� huonoja muutoksia. Immuunisysteemiss� on tapoja tunnistaa hyv�t ja huonot muutokset, Spetner kutstuu t�t� "hienos��d�ksi". Luonnossa vastaavaa ei kutsuta "hienos��d�ksi" vaan "luonnonvalinnaksi" - ja t�t� evoluutikot pit�v�t kovasti arvossa.
*Muutokset eiv�t tallennu perim��n.
Spetnerin k�sitys muuttuu t�ll� perusteella hieman kummalliseksi, koska h�nen v�itteens� on ymm�rrett�viss� "Informaatiota ei voi synty�, jos lopputulos tallennetaan." Lis�ongelmia t�lle tuo se, ett� vaikka onkin totta, ett� immuunisysteemi ei tallenna muutoksia ituradan perim��n, niin silti tiedot tallentuvat immuunisysteemin soluihin: syntyy resistenssi ja sama tauti paranee seuraavalla kerralla nopeammin. Ilmeisesti pelkk� tallennuskaan ei siis voi olla este, vaan ilmeisesti jokin vaikutus on sill� minne se tallennus tehd��n.. Tai sitten t�m� argumentti on n�enn�inen.
Kun n�m� kaikki kohdat otetaan huomioon on helpohko huomata, ett� immuunisysteemi on "analoginen" evoluutioprosesseihin. Evoluutiokin on hienostunut systeemi, jossa ymp�rist� asettaa rajaukset informaatiolle, ja jossa esiintyy runsaasti mutaatioita ja jossa on valintamekanismi.
Yhteenvetoa
"The NREH, on the other hand, postulates nonrandom variation. It does not fulfill the neo-Darwinian agenda in that it does not contribute to a natural explanation of the origin of life."
(Lee Spetner, "Not by Chance!", 1998, sivu 210)
Spetnerin keskittyy evoluutiokritiikkiin, mutta muista poiketen esitt�� my�s vastahypoteesin, "NREH":n.
*Evoluutiohypoteesissaan Spetner nojaa omaan informaatiom��rittelyyns�, joka ei ole kovin osuva -Spetner ei esimerkiksi k�sit�, ett� entsyymien ja substraattien sitoutumisella ei ole "kaikki tai ei mit��n" -luonnetta. H�n my�s laittaa yht�l�isyysmerkit "puhtaan Sattuman" ja "evoluution" v�liin. Spetnerin k�ytt�m� "hienos��t�perustelu", joka sallii informaation kasvun taas on ominaisuuksiltaan kovasti samanlainen, kuin luonnonvalinta.
*NREH:ssa keskeisint� on "evoluution ennalta ohjelmoitu potentiaali", joka m��r�� miten kehitys menee ja mink�laista informaatiota tulee. Spetner ei kuitenkaan selit� miten mutaatiot kaivavat potentiaalista informaatiota, eik� sit� miten t�t� potentiaalia voitaisiin tutkia. H�n paljastaa, ett� osa on "nukkuvia geenej�", mutta t�m� ei ole ainut menetelm�. Spetner ei my�sk��n kerro, syntyyk� "NREH":n piiriss� uutta informaatiota vai ei. Emme loppupeleiss� tied� mik� t�m� "potentiaali" on, emmek� my�sk��n mitk� ovat "NREH":n rajat, ja t�t� kautta kautta meill� ei ole keinoa falsifioida Spetnerin teoriaa: Esimerkiksi havainnot, joissa Spetnerin m��ritelm�n mukainen informaatio lis��ntyy ei falsifioi h�nen malliaan, koska ne voidaan tulkita "tuntemattomasta potentiaalista" johtuviksi. Spetner ei my�sk��n sovella evoluutioon soveltamaansa "Voisiko-se-toimia?" -kaavaansa omaan informaatiomalliinsa: l�hinn� koska h�nen mallinsa ei ainakaan toistaiseksi kerro mit��n sellaista mit� evoluutiokaan ei kertoisi - h�nen mallinsa keskeisimm�t, eli "mutaatioita ohjaava voima" ja "mikroevoluution potentiaali" kun ovat molemmat paitsi tuntemattomia my�s tuntemattoman suuruisia.
Kaikesta huolimatta Spetnerin malli ansaitsee erikoishuomion siit�, ett� se on toistaiseksi ainut kreationistien informaatiomalli, joka on sek� formalisaatio ett� vaihtoehtoinen malli.
Lis�tietoja:
Timo Tiainen,"Kriittinen arvio Spetnerin n�kemyksist�",2004
Ian Musgrave,"Information Theory and Creationism - Spetner and Biological Information",2005
Gert Korthof,"Could it work?",2003
Mark Chu-Carroll, "Using Bad Math to Create Bad Models to Produce Bad Results",2007
Oletetaan nyt ett� meill� on hyvyysfunktio jossa on lokaali optimi, joka ei kuitenkaan ole "korkein mahdollinen huippu". Sitten l�yd�mme organismin, joka kiipe�� siihen. Kun evoluutio saavuttaa maksimin, evoluutio lakkaa etenem�st�, jonka seurauksena juuri sen lokaalin optimin kohdalla alkaa olemaan ruuhkaa: Yksil�t kuluttavat samaa ravintoa, joka taas vaikuttaa hyvyysfunktioon. Seurauksena lokaalin optimin paikka muuttuu.
Lis�ksi evoluutio on moniulotteinen, joten pit�isi todistaa ett� optimi olisi rajattu joka suuntaan. Jos meill� on hyvyysavaruus, jolla on X ja Y akseli, ei voida sanoa ett� se ett� kohde on sellaisella kohdalla ett� sen Y on lokaalissa optimissa, se ei tarkoita ett� se ei voisi muuttua paremmaksi, se voi liikkua X:n kautta, ja t�m� X:n muutos voi jopa tarjota uuden kohdan, jossa Y ei en�� olekaan sidottu. (Eli lukkiutumia voi synty� ja ne voivat vaihtaa paikkaa.) "Lokaali optimi" ei siksi toimi "mikroevoluutioesteen� jota ei voi ylitt��".
Eli "hyvyysfunktiot" ovat tuskin koskaan staattisia ja "useampi ulottuvuus" tuo lis�ksi uusia kiertoteit�.
Toisin sanoen, Spetnerist� s��nn�nmukainen kehitys ei suinkaan osoita, ett� h�nen omat laskelmansa "rappeutumisen pakosta" ja "evoluution satunnaisuudesta eli kyvytt�myydest� tuottaa ennusteita", olisivat v��r�ss�. H�n ei my�sk��n p���ttele, ett� "informaation trendi on nouseva", vaan sen sijaan se kertoo siit�, ett� muuntelumahdollisuuksien m��r� on rajallinen ja n�ist� vaihtoehdoista valikoituu tietty "sattumanvaraisesti eli ymp�rist�st� riippuen". Samanaikaisesti, kun "aktuaalinen, havaittava informaatio" kasvaa, niin "potentiaali pienenee".
T�lt� kohdin on eritt�in vaikeaa n�hd� eroa evoluutikkojen n�kemykseen ; heist� luonnonvalinta valikoi hyv�t ja huonot ominaisuudet ja ett� "jossain on potentiaalisessa muodossa tieto siit� millaista informaatiota eli�iss� voi olla". -Barry Hallin lis�ksi mm. Richard Dawkins on kirjoittanut luonnonvalinnan tuottamasta ennustettavuudesta- Evoluutikoista universumin sis�lt�m�t luonnonlait sek� n�iden keskin�inen vaikutus ymp�rist�ss� sis�lt�v�t "potentiaalisessa muodossa" tiedon "mink�lainen eli� on siihen sopiva ja mik� v�hemm�n sopiva". Spetner tulkitsee osan informaatiosta "nukkuviksi geeneiksi", mutta ei rajaa t�t� ainoaksi vaihtoehdoksi. Sill� miss� "potentiaali -informaation" varasto on, ei ole merkitt�v� asia. "Mikroevoluution potentiaali" voidaan siis varsin hyvin ymm�rt�� my�s "teistiseksi evoluutioksi" eik� oikein mitenk��n "evoluutiokritiikiksi".
Toisin sanoen Spetner ei vetoa ulkoisiin rakenteisiin. H�nen n�kemyksens� kannalta on t�ysin mahdollista, ett� koko eli�kunta olisi l�ht�isin esimerkiksi pienen bakteerin "potentiaalista". T�m� tarkoittaa sit�, ett� Spetner voi ajatella nykyisest� fossiilirekordista sen, ett� "potentiaali -informaatio" selv�sti ajan kanssa muuttuu "aktuaaliseksi informaatioksi".
T�ll� n�kemyksell� on muutamia heikkouksia:
*Ensinn�kin mink�lainen tahansa fossiiliaineisto sopii Spetnerin n�kemykseen, eli fossiileilla n�kemyst� ei voi falsifioida.
*Toisaalta t�m� johtaa mu�s siihen, ett� "rappeutuneiden rakenteiden" esittely ei my�sk��n ole mik��n todiste : Informaatiom��r� kun ei ole pelk�st��n n�kyviss� rakenteissa.
*Kolmanneksi jos potentiaali olisi koodattuna perim��n, ja se vain "nousisi sielt� ajan mittaan esiin", pit�isi mallin selitt�� miten; Ongelmana kun on se, ett� eitoiminnallinen, piilossa oleva, potentiaali on kuitenkin altis satunnaisille mutaatioille, jonka pit�isi johtaa siihen ett� ilman valintapainetta potentiaali tuhoutuu pitk�ll� t�ht�imell�: Toisin sanoen siin� miss� evoluutio EI v�it� ett� ominaisuudet nousisivat esiin sattumalta, vaan p�invastoin korostavat luonnonvalinnan merkityst� siin� ett� toiminta ei tuhoudu, koodattuun, eitoiminnalliseen, potentiaaliin uskova joutuisi hyv�ksym��n "sattumalta esiin nousevan informaation". Potentiaali voisi toki nousta esiin "eisatunnaisin mutaatioin", mutta t�ll�isest� manipulaatiosta ei ole havaintoja.
Spetnerin malli koskee pelk�st��n entsyymeit�, eik� siksi ole sovellettavissa rakenteisiin tai mihink��n muuhun. Spetnerin mallin falsifiointi on kyseenalaista, koska periaatteessa mik� tahansa informaatio voi olla per�isin "jostakin tuntemattomasta potentiaalista" ja vaikka jokin mutaatio muuttaisi jotain entsyymi� "spesifisemm�ksi", on se ainoastaan "spesifisemm�ksi suhteessa tiet�miimme substraatteihin", jolloin ei voida tiet�� onko kyse "aidosta spesifioitumisesta" eli "spesifisyydest� suhteessa kaikkiin substraatteihin". Jos potentiaali katsotaan rajoitetuksi pelk�st��n perim��n, niin tilanne muuttuu. Spetner muuttuu empiriaksi heti, kun tapa mitata potentiaalin suuruus keksit��n, ja heti, kun Spetner onnistuu keksim��n sen mekanismin, joka takaa sen, ett� mutaatiot ovatkin hy�dyllisi�, informaatiota "potentiaalista" esiinkaivavia, sen sijaan ett� olisivat "satunnaisia".
Mutaatiot - uuden rakentaminen vai varastostahaku
"I agree with him that these mutations add information to the B-cell genome."
(Lee Spetner, "Lee Spetner/Edward Max Dialogue", 2006)
On havaittu sellaisia "satunnaisia" mutaatioita, jotka lis��v�t geneettist� informaatiota jopa Spetnerin menetelm�ll� mitattuna. Erityisen vaikeaksi Spetnerin mallille on ollut immuunisysteemi, koska vasta -aineet eiv�t ole yhdistettyn� "nukkuviin geeneihin", vaan ensimm�isell� kerralla kun tauti iskee, siihen sopivaa informaatiota ei ole, vaan se on generoitava satunnaisin mutaatioin. Spetner on my�nt�nyt sen, ett� immuunisysteemi lis�� informaatiota B -solun genomiin. Kyse ei siis ole "potentiaali - informaation esiinkaivamisesta", vaan aidosta informaation kasvusta. Eli jotain, jonka piti h�nen omista m��ritelmist��n k�sin olla mahdotonta.
Spetner esitt�� seuraavia syit� sille,miksi t�m� on mahdollista:
*Mutaatiom��r� on suuri : Arviot v�lill� 100 miljoonaa - 10 miljardia kappaletta.
T�m�n p��telm�n Spetner perustelee, sivulla 103, sill� ett� todenn�k�isyys tuottaa informaatiota "evoluutiosattumalta" saadaan, kun kerrotaan jokaisen v�livaiheen todenn�k�isyydet kesken��n : T�m� todenn�k�isyys taas saavutetaan arvaamalla tarpeeksi monta kertaa. T�ss� n�kemyksess� kummallista on:
-Luonnonvalintaa ei oteta huomioon lainkaan, jolloin v�livaiheiden k�ytt� itse asiassa menett�� merkityksens�. V�livaiheella voi olla merkityst� vain luonnonvalinnan kautta. "V�livaiheilla" luodaan ainoastaan valheellinen mielikuva "evoluution huomioon ottamisessa".
-Miten t�t� selityst� soveltaen informaatiota ei synny luonnossa "miljoonissa vuosissa", kun todenn�k�isyydet pysyv�t ja mutaatiot tuottavat jatkuvasti "arvauksia"? Eik� sen pit�isi olla pikemminkin v�ist�m�t�nt�?
On joka tapauksessa eritt�in erikoista, ett� Spetner, jotka kiist�� "aidosti hy�dyllisten mutaatioiden" olemassaolonkin kaikesta huolimatta itse tarjoaa mutaatioita "tarpeeksi isoissa m��riss� esiintyess��n" selityksen� "aidosti uuden informaation" synnylle. Eih�n "puhtaan todenn�k�isyyden" t�yttymiselle, sellaisenahan Spetner tilanteen laskee- ole ratkaisevaa se, kuinka lyhyess� ajassa yritykset tapahtuvat, vaan siin� kuinka monta kertaa n�in tapahtuu.
*Immuunisysteemiss� k�sitelt�v� informaatiom��r� on pieni
T�m� on "virhekatastrofirajaan" sovellettuna aivan j�rkev� argumentti. Suuret mutaatiom��r�t kuitenkin toimisivat hajottavana voimana. Perim�ss�mme mutaatiom��r� on selv�sti immuunisysteemi� pienempi informaatiom��r� ovat selv�sti suurempi. N�in suurempi informaatiom��r� joudutaan kompensoimaan, eik� virhekatastrofiin ajauduta.
*Systeemi on hienos��detty.
On aivan totta, ett� informaatio "on biteiss�": sek� informaation kasvu ett� sen v�heneminen ovat muutoksia niiss� - luokiteltiin informaatio mill� tavoin tahansa. N�in ollen pelk�t muutokset eiv�t riit�, vaan tarvitaan jotain, joka "tarkastaa" mitk� ovat hyvi� ja mitk� huonoja muutoksia. Immuunisysteemiss� on tapoja tunnistaa hyv�t ja huonot muutokset, Spetner kutstuu t�t� "hienos��d�ksi". Luonnossa vastaavaa ei kutsuta "hienos��d�ksi" vaan "luonnonvalinnaksi" - ja t�t� evoluutikot pit�v�t kovasti arvossa.
*Muutokset eiv�t tallennu perim��n.
Spetnerin k�sitys muuttuu t�ll� perusteella hieman kummalliseksi, koska h�nen v�itteens� on ymm�rrett�viss� "Informaatiota ei voi synty�, jos lopputulos tallennetaan." Lis�ongelmia t�lle tuo se, ett� vaikka onkin totta, ett� immuunisysteemi ei tallenna muutoksia ituradan perim��n, niin silti tiedot tallentuvat immuunisysteemin soluihin: syntyy resistenssi ja sama tauti paranee seuraavalla kerralla nopeammin. Ilmeisesti pelkk� tallennuskaan ei siis voi olla este, vaan ilmeisesti jokin vaikutus on sill� minne se tallennus tehd��n.. Tai sitten t�m� argumentti on n�enn�inen.
Kun n�m� kaikki kohdat otetaan huomioon on helpohko huomata, ett� immuunisysteemi on "analoginen" evoluutioprosesseihin. Evoluutiokin on hienostunut systeemi, jossa ymp�rist� asettaa rajaukset informaatiolle, ja jossa esiintyy runsaasti mutaatioita ja jossa on valintamekanismi.
Yhteenvetoa
"The NREH, on the other hand, postulates nonrandom variation. It does not fulfill the neo-Darwinian agenda in that it does not contribute to a natural explanation of the origin of life."
(Lee Spetner, "Not by Chance!", 1998, sivu 210)
Spetnerin keskittyy evoluutiokritiikkiin, mutta muista poiketen esitt�� my�s vastahypoteesin, "NREH":n.
*Evoluutiohypoteesissaan Spetner nojaa omaan informaatiom��rittelyyns�, joka ei ole kovin osuva -Spetner ei esimerkiksi k�sit�, ett� entsyymien ja substraattien sitoutumisella ei ole "kaikki tai ei mit��n" -luonnetta. H�n my�s laittaa yht�l�isyysmerkit "puhtaan Sattuman" ja "evoluution" v�liin. Spetnerin k�ytt�m� "hienos��t�perustelu", joka sallii informaation kasvun taas on ominaisuuksiltaan kovasti samanlainen, kuin luonnonvalinta.
*NREH:ssa keskeisint� on "evoluution ennalta ohjelmoitu potentiaali", joka m��r�� miten kehitys menee ja mink�laista informaatiota tulee. Spetner ei kuitenkaan selit� miten mutaatiot kaivavat potentiaalista informaatiota, eik� sit� miten t�t� potentiaalia voitaisiin tutkia. H�n paljastaa, ett� osa on "nukkuvia geenej�", mutta t�m� ei ole ainut menetelm�. Spetner ei my�sk��n kerro, syntyyk� "NREH":n piiriss� uutta informaatiota vai ei. Emme loppupeleiss� tied� mik� t�m� "potentiaali" on, emmek� my�sk��n mitk� ovat "NREH":n rajat, ja t�t� kautta kautta meill� ei ole keinoa falsifioida Spetnerin teoriaa: Esimerkiksi havainnot, joissa Spetnerin m��ritelm�n mukainen informaatio lis��ntyy ei falsifioi h�nen malliaan, koska ne voidaan tulkita "tuntemattomasta potentiaalista" johtuviksi. Spetner ei my�sk��n sovella evoluutioon soveltamaansa "Voisiko-se-toimia?" -kaavaansa omaan informaatiomalliinsa: l�hinn� koska h�nen mallinsa ei ainakaan toistaiseksi kerro mit��n sellaista mit� evoluutiokaan ei kertoisi - h�nen mallinsa keskeisimm�t, eli "mutaatioita ohjaava voima" ja "mikroevoluution potentiaali" kun ovat molemmat paitsi tuntemattomia my�s tuntemattoman suuruisia.
Kaikesta huolimatta Spetnerin malli ansaitsee erikoishuomion siit�, ett� se on toistaiseksi ainut kreationistien informaatiomalli, joka on sek� formalisaatio ett� vaihtoehtoinen malli.
Lis�tietoja:
Timo Tiainen,"Kriittinen arvio Spetnerin n�kemyksist�",2004
Ian Musgrave,"Information Theory and Creationism - Spetner and Biological Information",2005
Gert Korthof,"Could it work?",2003
Mark Chu-Carroll, "Using Bad Math to Create Bad Models to Produce Bad Results",2007
Toisin sanoen Spetner ei vetoa ulkoisiin rakenteisiin. H�nen n�kemyksens� kannalta on t�ysin mahdollista, ett� koko eli�kunta olisi l�ht�isin esimerkiksi pienen bakteerin "potentiaalista". T�m� tarkoittaa sit�, ett� Spetner voi ajatella nykyisest� fossiilirekordista sen, ett� "potentiaali -informaatio" selv�sti ajan kanssa muuttuu "aktuaaliseksi informaatioksi".
T�ll� n�kemyksell� on muutamia heikkouksia:
*Ensinn�kin mink�lainen tahansa fossiiliaineisto sopii Spetnerin n�kemykseen, eli fossiileilla n�kemyst� ei voi falsifioida.
*Toisaalta t�m� johtaa mu�s siihen, ett� "rappeutuneiden rakenteiden" esittely ei my�sk��n ole mik��n todiste : Informaatiom��r� kun ei ole pelk�st��n n�kyviss� rakenteissa.
*Kolmanneksi jos potentiaali olisi koodattuna perim��n, ja se vain "nousisi sielt� ajan mittaan esiin", pit�isi mallin selitt�� miten; Ongelmana kun on se, ett� eitoiminnallinen, piilossa oleva, potentiaali on kuitenkin altis satunnaisille mutaatioille, jonka pit�isi johtaa siihen ett� ilman valintapainetta potentiaali tuhoutuu pitk�ll� t�ht�imell�: Toisin sanoen siin� miss� evoluutio EI v�it� ett� ominaisuudet nousisivat esiin sattumalta, vaan p�invastoin korostavat luonnonvalinnan merkityst� siin� ett� toiminta ei tuhoudu, koodattuun, eitoiminnalliseen, potentiaaliin uskova joutuisi hyv�ksym��n "sattumalta esiin nousevan informaation". Potentiaali voisi toki nousta esiin "eisatunnaisin mutaatioin", mutta t�ll�isest� manipulaatiosta ei ole havaintoja.
Spetnerin malli koskee pelk�st��n entsyymeit�, eik� siksi ole sovellettavissa rakenteisiin tai mihink��n muuhun. Spetnerin mallin falsifiointi on kyseenalaista, koska periaatteessa mik� tahansa informaatio voi olla per�isin "jostakin tuntemattomasta potentiaalista" ja vaikka jokin mutaatio muuttaisi jotain entsyymi� "spesifisemm�ksi", on se ainoastaan "spesifisemm�ksi suhteessa tiet�miimme substraatteihin", jolloin ei voida tiet�� onko kyse "aidosta spesifioitumisesta" eli "spesifisyydest� suhteessa kaikkiin substraatteihin". Jos potentiaali katsotaan rajoitetuksi pelk�st��n perim��n, niin tilanne muuttuu. Spetner muuttuu empiriaksi heti, kun tapa mitata potentiaalin suuruus keksit��n, ja heti, kun Spetner onnistuu keksim��n sen mekanismin, joka takaa sen, ett� mutaatiot ovatkin hy�dyllisi�, informaatiota "potentiaalista" esiinkaivavia, sen sijaan ett� olisivat "satunnaisia".
Mutaatiot - uuden rakentaminen vai varastostahaku
"I agree with him that these mutations add information to the B-cell genome."
(Lee Spetner, "Lee Spetner/Edward Max Dialogue", 2006)
On havaittu sellaisia "satunnaisia" mutaatioita, jotka lis��v�t geneettist� informaatiota jopa Spetnerin menetelm�ll� mitattuna. Erityisen vaikeaksi Spetnerin mallille on ollut immuunisysteemi, koska vasta -aineet eiv�t ole yhdistettyn� "nukkuviin geeneihin", vaan ensimm�isell� kerralla kun tauti iskee, siihen sopivaa informaatiota ei ole, vaan se on generoitava satunnaisin mutaatioin. Spetner on my�nt�nyt sen, ett� immuunisysteemi lis�� informaatiota B -solun genomiin. Kyse ei siis ole "potentiaali - informaation esiinkaivamisesta", vaan aidosta informaation kasvusta. Eli jotain, jonka piti h�nen omista m��ritelmist��n k�sin olla mahdotonta.
Spetner esitt�� seuraavia syit� sille,miksi t�m� on mahdollista:
*Mutaatiom��r� on suuri : Arviot v�lill� 100 miljoonaa - 10 miljardia kappaletta.
T�m�n p��telm�n Spetner perustelee, sivulla 103, sill� ett� todenn�k�isyys tuottaa informaatiota "evoluutiosattumalta" saadaan, kun kerrotaan jokaisen v�livaiheen todenn�k�isyydet kesken��n : T�m� todenn�k�isyys taas saavutetaan arvaamalla tarpeeksi monta kertaa. T�ss� n�kemyksess� kummallista on:
-Luonnonvalintaa ei oteta huomioon lainkaan, jolloin v�livaiheiden k�ytt� itse asiassa menett�� merkityksens�. V�livaiheella voi olla merkityst� vain luonnonvalinnan kautta. "V�livaiheilla" luodaan ainoastaan valheellinen mielikuva "evoluution huomioon ottamisessa".
-Miten t�t� selityst� soveltaen informaatiota ei synny luonnossa "miljoonissa vuosissa", kun todenn�k�isyydet pysyv�t ja mutaatiot tuottavat jatkuvasti "arvauksia"? Eik� sen pit�isi olla pikemminkin v�ist�m�t�nt�?
On joka tapauksessa eritt�in erikoista, ett� Spetner, jotka kiist�� "aidosti hy�dyllisten mutaatioiden" olemassaolonkin kaikesta huolimatta itse tarjoaa mutaatioita "tarpeeksi isoissa m��riss� esiintyess��n" selityksen� "aidosti uuden informaation" synnylle. Eih�n "puhtaan todenn�k�isyyden" t�yttymiselle, sellaisenahan Spetner tilanteen laskee- ole ratkaisevaa se, kuinka lyhyess� ajassa yritykset tapahtuvat, vaan siin� kuinka monta kertaa n�in tapahtuu.
*Immuunisysteemiss� k�sitelt�v� informaatiom��r� on pieni
T�m� on "virhekatastrofirajaan" sovellettuna aivan j�rkev� argumentti. Suuret mutaatiom��r�t kuitenkin toimisivat hajottavana voimana. Perim�ss�mme mutaatiom��r� on selv�sti immuunisysteemi� pienempi informaatiom��r� ovat selv�sti suurempi. N�in suurempi informaatiom��r� joudutaan kompensoimaan, eik� virhekatastrofiin ajauduta.
*Systeemi on hienos��detty.
On aivan totta, ett� informaatio "on biteiss�": sek� informaation kasvu ett� sen v�heneminen ovat muutoksia niiss� - luokiteltiin informaatio mill� tavoin tahansa. N�in ollen pelk�t muutokset eiv�t riit�, vaan tarvitaan jotain, joka "tarkastaa" mitk� ovat hyvi� ja mitk� huonoja muutoksia. Immuunisysteemiss� on tapoja tunnistaa hyv�t ja huonot muutokset, Spetner kutstuu t�t� "hienos��d�ksi". Luonnossa vastaavaa ei kutsuta "hienos��d�ksi" vaan "luonnonvalinnaksi" - ja t�t� evoluutikot pit�v�t kovasti arvossa.
*Muutokset eiv�t tallennu perim��n.
Spetnerin k�sitys muuttuu t�ll� perusteella hieman kummalliseksi, koska h�nen v�itteens� on ymm�rrett�viss� "Informaatiota ei voi synty�, jos lopputulos tallennetaan." Lis�ongelmia t�lle tuo se, ett� vaikka onkin totta, ett� immuunisysteemi ei tallenna muutoksia ituradan perim��n, niin silti tiedot tallentuvat immuunisysteemin soluihin: syntyy resistenssi ja sama tauti paranee seuraavalla kerralla nopeammin. Ilmeisesti pelkk� tallennuskaan ei siis voi olla este, vaan ilmeisesti jokin vaikutus on sill� minne se tallennus tehd��n.. Tai sitten t�m� argumentti on n�enn�inen.
Kun n�m� kaikki kohdat otetaan huomioon on helpohko huomata, ett� immuunisysteemi on "analoginen" evoluutioprosesseihin. Evoluutiokin on hienostunut systeemi, jossa ymp�rist� asettaa rajaukset informaatiolle, ja jossa esiintyy runsaasti mutaatioita ja jossa on valintamekanismi.
Yhteenvetoa
"The NREH, on the other hand, postulates nonrandom variation. It does not fulfill the neo-Darwinian agenda in that it does not contribute to a natural explanation of the origin of life."
(Lee Spetner, "Not by Chance!", 1998, sivu 210)
Spetnerin keskittyy evoluutiokritiikkiin, mutta muista poiketen esitt�� my�s vastahypoteesin, "NREH":n.
*Evoluutiohypoteesissaan Spetner nojaa omaan informaatiom��rittelyyns�, joka ei ole kovin osuva -Spetner ei esimerkiksi k�sit�, ett� entsyymien ja substraattien sitoutumisella ei ole "kaikki tai ei mit��n" -luonnetta. H�n my�s laittaa yht�l�isyysmerkit "puhtaan Sattuman" ja "evoluution" v�liin. Spetnerin k�ytt�m� "hienos��t�perustelu", joka sallii informaation kasvun taas on ominaisuuksiltaan kovasti samanlainen, kuin luonnonvalinta.
*NREH:ssa keskeisint� on "evoluution ennalta ohjelmoitu potentiaali", joka m��r�� miten kehitys menee ja mink�laista informaatiota tulee. Spetner ei kuitenkaan selit� miten mutaatiot kaivavat potentiaalista informaatiota, eik� sit� miten t�t� potentiaalia voitaisiin tutkia. H�n paljastaa, ett� osa on "nukkuvia geenej�", mutta t�m� ei ole ainut menetelm�. Spetner ei my�sk��n kerro, syntyyk� "NREH":n piiriss� uutta informaatiota vai ei. Emme loppupeleiss� tied� mik� t�m� "potentiaali" on, emmek� my�sk��n mitk� ovat "NREH":n rajat, ja t�t� kautta kautta meill� ei ole keinoa falsifioida Spetnerin teoriaa: Esimerkiksi havainnot, joissa Spetnerin m��ritelm�n mukainen informaatio lis��ntyy ei falsifioi h�nen malliaan, koska ne voidaan tulkita "tuntemattomasta potentiaalista" johtuviksi. Spetner ei my�sk��n sovella evoluutioon soveltamaansa "Voisiko-se-toimia?" -kaavaansa omaan informaatiomalliinsa: l�hinn� koska h�nen mallinsa ei ainakaan toistaiseksi kerro mit��n sellaista mit� evoluutiokaan ei kertoisi - h�nen mallinsa keskeisimm�t, eli "mutaatioita ohjaava voima" ja "mikroevoluution potentiaali" kun ovat molemmat paitsi tuntemattomia my�s tuntemattoman suuruisia.
Kaikesta huolimatta Spetnerin malli ansaitsee erikoishuomion siit�, ett� se on toistaiseksi ainut kreationistien informaatiomalli, joka on sek� formalisaatio ett� vaihtoehtoinen malli.
Lis�tietoja:
Timo Tiainen,"Kriittinen arvio Spetnerin n�kemyksist�",2004
Ian Musgrave,"Information Theory and Creationism - Spetner and Biological Information",2005
Gert Korthof,"Could it work?",2003
Mark Chu-Carroll, "Using Bad Math to Create Bad Models to Produce Bad Results",2007
"I agree with him that these mutations add information to the B-cell genome."
(Lee Spetner, "Lee Spetner/Edward Max Dialogue", 2006)
On havaittu sellaisia "satunnaisia" mutaatioita, jotka lis��v�t geneettist� informaatiota jopa Spetnerin menetelm�ll� mitattuna. Erityisen vaikeaksi Spetnerin mallille on ollut immuunisysteemi, koska vasta -aineet eiv�t ole yhdistettyn� "nukkuviin geeneihin", vaan ensimm�isell� kerralla kun tauti iskee, siihen sopivaa informaatiota ei ole, vaan se on generoitava satunnaisin mutaatioin. Spetner on my�nt�nyt sen, ett� immuunisysteemi lis�� informaatiota B -solun genomiin. Kyse ei siis ole "potentiaali - informaation esiinkaivamisesta", vaan aidosta informaation kasvusta. Eli jotain, jonka piti h�nen omista m��ritelmist��n k�sin olla mahdotonta.
Spetner esitt�� seuraavia syit� sille,miksi t�m� on mahdollista:
*Mutaatiom��r� on suuri : Arviot v�lill� 100 miljoonaa - 10 miljardia kappaletta.
T�m�n p��telm�n Spetner perustelee, sivulla 103, sill� ett� todenn�k�isyys tuottaa informaatiota "evoluutiosattumalta" saadaan, kun kerrotaan jokaisen v�livaiheen todenn�k�isyydet kesken��n : T�m� todenn�k�isyys taas saavutetaan arvaamalla tarpeeksi monta kertaa. T�ss� n�kemyksess� kummallista on:
-Luonnonvalintaa ei oteta huomioon lainkaan, jolloin v�livaiheiden k�ytt� itse asiassa menett�� merkityksens�. V�livaiheella voi olla merkityst� vain luonnonvalinnan kautta. "V�livaiheilla" luodaan ainoastaan valheellinen mielikuva "evoluution huomioon ottamisessa".
-Miten t�t� selityst� soveltaen informaatiota ei synny luonnossa "miljoonissa vuosissa", kun todenn�k�isyydet pysyv�t ja mutaatiot tuottavat jatkuvasti "arvauksia"? Eik� sen pit�isi olla pikemminkin v�ist�m�t�nt�?
On joka tapauksessa eritt�in erikoista, ett� Spetner, jotka kiist�� "aidosti hy�dyllisten mutaatioiden" olemassaolonkin kaikesta huolimatta itse tarjoaa mutaatioita "tarpeeksi isoissa m��riss� esiintyess��n" selityksen� "aidosti uuden informaation" synnylle. Eih�n "puhtaan todenn�k�isyyden" t�yttymiselle, sellaisenahan Spetner tilanteen laskee- ole ratkaisevaa se, kuinka lyhyess� ajassa yritykset tapahtuvat, vaan siin� kuinka monta kertaa n�in tapahtuu.
*Immuunisysteemiss� k�sitelt�v� informaatiom��r� on pieni
T�m� on "virhekatastrofirajaan" sovellettuna aivan j�rkev� argumentti. Suuret mutaatiom��r�t kuitenkin toimisivat hajottavana voimana. Perim�ss�mme mutaatiom��r� on selv�sti immuunisysteemi� pienempi informaatiom��r� ovat selv�sti suurempi. N�in suurempi informaatiom��r� joudutaan kompensoimaan, eik� virhekatastrofiin ajauduta.
*Systeemi on hienos��detty.
On aivan totta, ett� informaatio "on biteiss�": sek� informaation kasvu ett� sen v�heneminen ovat muutoksia niiss� - luokiteltiin informaatio mill� tavoin tahansa. N�in ollen pelk�t muutokset eiv�t riit�, vaan tarvitaan jotain, joka "tarkastaa" mitk� ovat hyvi� ja mitk� huonoja muutoksia. Immuunisysteemiss� on tapoja tunnistaa hyv�t ja huonot muutokset, Spetner kutstuu t�t� "hienos��d�ksi". Luonnossa vastaavaa ei kutsuta "hienos��d�ksi" vaan "luonnonvalinnaksi" - ja t�t� evoluutikot pit�v�t kovasti arvossa.
*Muutokset eiv�t tallennu perim��n.
Spetnerin k�sitys muuttuu t�ll� perusteella hieman kummalliseksi, koska h�nen v�itteens� on ymm�rrett�viss� "Informaatiota ei voi synty�, jos lopputulos tallennetaan." Lis�ongelmia t�lle tuo se, ett� vaikka onkin totta, ett� immuunisysteemi ei tallenna muutoksia ituradan perim��n, niin silti tiedot tallentuvat immuunisysteemin soluihin: syntyy resistenssi ja sama tauti paranee seuraavalla kerralla nopeammin. Ilmeisesti pelkk� tallennuskaan ei siis voi olla este, vaan ilmeisesti jokin vaikutus on sill� minne se tallennus tehd��n.. Tai sitten t�m� argumentti on n�enn�inen.
Kun n�m� kaikki kohdat otetaan huomioon on helpohko huomata, ett� immuunisysteemi on "analoginen" evoluutioprosesseihin. Evoluutiokin on hienostunut systeemi, jossa ymp�rist� asettaa rajaukset informaatiolle, ja jossa esiintyy runsaasti mutaatioita ja jossa on valintamekanismi.
Yhteenvetoa
"The NREH, on the other hand, postulates nonrandom variation. It does not fulfill the neo-Darwinian agenda in that it does not contribute to a natural explanation of the origin of life."
(Lee Spetner, "Not by Chance!", 1998, sivu 210)
Spetnerin keskittyy evoluutiokritiikkiin, mutta muista poiketen esitt�� my�s vastahypoteesin, "NREH":n.
*Evoluutiohypoteesissaan Spetner nojaa omaan informaatiom��rittelyyns�, joka ei ole kovin osuva -Spetner ei esimerkiksi k�sit�, ett� entsyymien ja substraattien sitoutumisella ei ole "kaikki tai ei mit��n" -luonnetta. H�n my�s laittaa yht�l�isyysmerkit "puhtaan Sattuman" ja "evoluution" v�liin. Spetnerin k�ytt�m� "hienos��t�perustelu", joka sallii informaation kasvun taas on ominaisuuksiltaan kovasti samanlainen, kuin luonnonvalinta.
*NREH:ssa keskeisint� on "evoluution ennalta ohjelmoitu potentiaali", joka m��r�� miten kehitys menee ja mink�laista informaatiota tulee. Spetner ei kuitenkaan selit� miten mutaatiot kaivavat potentiaalista informaatiota, eik� sit� miten t�t� potentiaalia voitaisiin tutkia. H�n paljastaa, ett� osa on "nukkuvia geenej�", mutta t�m� ei ole ainut menetelm�. Spetner ei my�sk��n kerro, syntyyk� "NREH":n piiriss� uutta informaatiota vai ei. Emme loppupeleiss� tied� mik� t�m� "potentiaali" on, emmek� my�sk��n mitk� ovat "NREH":n rajat, ja t�t� kautta kautta meill� ei ole keinoa falsifioida Spetnerin teoriaa: Esimerkiksi havainnot, joissa Spetnerin m��ritelm�n mukainen informaatio lis��ntyy ei falsifioi h�nen malliaan, koska ne voidaan tulkita "tuntemattomasta potentiaalista" johtuviksi. Spetner ei my�sk��n sovella evoluutioon soveltamaansa "Voisiko-se-toimia?" -kaavaansa omaan informaatiomalliinsa: l�hinn� koska h�nen mallinsa ei ainakaan toistaiseksi kerro mit��n sellaista mit� evoluutiokaan ei kertoisi - h�nen mallinsa keskeisimm�t, eli "mutaatioita ohjaava voima" ja "mikroevoluution potentiaali" kun ovat molemmat paitsi tuntemattomia my�s tuntemattoman suuruisia.
Kaikesta huolimatta Spetnerin malli ansaitsee erikoishuomion siit�, ett� se on toistaiseksi ainut kreationistien informaatiomalli, joka on sek� formalisaatio ett� vaihtoehtoinen malli.
Lis�tietoja:
Timo Tiainen,"Kriittinen arvio Spetnerin n�kemyksist�",2004
Ian Musgrave,"Information Theory and Creationism - Spetner and Biological Information",2005
Gert Korthof,"Could it work?",2003
Mark Chu-Carroll, "Using Bad Math to Create Bad Models to Produce Bad Results",2007
Spetner esitt�� seuraavia syit� sille,miksi t�m� on mahdollista:
*Mutaatiom��r� on suuri : Arviot v�lill� 100 miljoonaa - 10 miljardia kappaletta.
T�m�n p��telm�n Spetner perustelee, sivulla 103, sill� ett� todenn�k�isyys tuottaa informaatiota "evoluutiosattumalta" saadaan, kun kerrotaan jokaisen v�livaiheen todenn�k�isyydet kesken��n : T�m� todenn�k�isyys taas saavutetaan arvaamalla tarpeeksi monta kertaa. T�ss� n�kemyksess� kummallista on:
-Luonnonvalintaa ei oteta huomioon lainkaan, jolloin v�livaiheiden k�ytt� itse asiassa menett�� merkityksens�. V�livaiheella voi olla merkityst� vain luonnonvalinnan kautta. "V�livaiheilla" luodaan ainoastaan valheellinen mielikuva "evoluution huomioon ottamisessa".
-Miten t�t� selityst� soveltaen informaatiota ei synny luonnossa "miljoonissa vuosissa", kun todenn�k�isyydet pysyv�t ja mutaatiot tuottavat jatkuvasti "arvauksia"? Eik� sen pit�isi olla pikemminkin v�ist�m�t�nt�?
On joka tapauksessa eritt�in erikoista, ett� Spetner, jotka kiist�� "aidosti hy�dyllisten mutaatioiden" olemassaolonkin kaikesta huolimatta itse tarjoaa mutaatioita "tarpeeksi isoissa m��riss� esiintyess��n" selityksen� "aidosti uuden informaation" synnylle. Eih�n "puhtaan todenn�k�isyyden" t�yttymiselle, sellaisenahan Spetner tilanteen laskee- ole ratkaisevaa se, kuinka lyhyess� ajassa yritykset tapahtuvat, vaan siin� kuinka monta kertaa n�in tapahtuu.
*Immuunisysteemiss� k�sitelt�v� informaatiom��r� on pieni
T�m� on "virhekatastrofirajaan" sovellettuna aivan j�rkev� argumentti. Suuret mutaatiom��r�t kuitenkin toimisivat hajottavana voimana. Perim�ss�mme mutaatiom��r� on selv�sti immuunisysteemi� pienempi informaatiom��r� ovat selv�sti suurempi. N�in suurempi informaatiom��r� joudutaan kompensoimaan, eik� virhekatastrofiin ajauduta.
*Systeemi on hienos��detty.
On aivan totta, ett� informaatio "on biteiss�": sek� informaation kasvu ett� sen v�heneminen ovat muutoksia niiss� - luokiteltiin informaatio mill� tavoin tahansa. N�in ollen pelk�t muutokset eiv�t riit�, vaan tarvitaan jotain, joka "tarkastaa" mitk� ovat hyvi� ja mitk� huonoja muutoksia. Immuunisysteemiss� on tapoja tunnistaa hyv�t ja huonot muutokset, Spetner kutstuu t�t� "hienos��d�ksi". Luonnossa vastaavaa ei kutsuta "hienos��d�ksi" vaan "luonnonvalinnaksi" - ja t�t� evoluutikot pit�v�t kovasti arvossa.
*Muutokset eiv�t tallennu perim��n.
Spetnerin k�sitys muuttuu t�ll� perusteella hieman kummalliseksi, koska h�nen v�itteens� on ymm�rrett�viss� "Informaatiota ei voi synty�, jos lopputulos tallennetaan." Lis�ongelmia t�lle tuo se, ett� vaikka onkin totta, ett� immuunisysteemi ei tallenna muutoksia ituradan perim��n, niin silti tiedot tallentuvat immuunisysteemin soluihin: syntyy resistenssi ja sama tauti paranee seuraavalla kerralla nopeammin. Ilmeisesti pelkk� tallennuskaan ei siis voi olla este, vaan ilmeisesti jokin vaikutus on sill� minne se tallennus tehd��n.. Tai sitten t�m� argumentti on n�enn�inen.
Kun n�m� kaikki kohdat otetaan huomioon on helpohko huomata, ett� immuunisysteemi on "analoginen" evoluutioprosesseihin. Evoluutiokin on hienostunut systeemi, jossa ymp�rist� asettaa rajaukset informaatiolle, ja jossa esiintyy runsaasti mutaatioita ja jossa on valintamekanismi.
Yhteenvetoa
"The NREH, on the other hand, postulates nonrandom variation. It does not fulfill the neo-Darwinian agenda in that it does not contribute to a natural explanation of the origin of life."
(Lee Spetner, "Not by Chance!", 1998, sivu 210)
Spetnerin keskittyy evoluutiokritiikkiin, mutta muista poiketen esitt�� my�s vastahypoteesin, "NREH":n.
*Evoluutiohypoteesissaan Spetner nojaa omaan informaatiom��rittelyyns�, joka ei ole kovin osuva -Spetner ei esimerkiksi k�sit�, ett� entsyymien ja substraattien sitoutumisella ei ole "kaikki tai ei mit��n" -luonnetta. H�n my�s laittaa yht�l�isyysmerkit "puhtaan Sattuman" ja "evoluution" v�liin. Spetnerin k�ytt�m� "hienos��t�perustelu", joka sallii informaation kasvun taas on ominaisuuksiltaan kovasti samanlainen, kuin luonnonvalinta.
*NREH:ssa keskeisint� on "evoluution ennalta ohjelmoitu potentiaali", joka m��r�� miten kehitys menee ja mink�laista informaatiota tulee. Spetner ei kuitenkaan selit� miten mutaatiot kaivavat potentiaalista informaatiota, eik� sit� miten t�t� potentiaalia voitaisiin tutkia. H�n paljastaa, ett� osa on "nukkuvia geenej�", mutta t�m� ei ole ainut menetelm�. Spetner ei my�sk��n kerro, syntyyk� "NREH":n piiriss� uutta informaatiota vai ei. Emme loppupeleiss� tied� mik� t�m� "potentiaali" on, emmek� my�sk��n mitk� ovat "NREH":n rajat, ja t�t� kautta kautta meill� ei ole keinoa falsifioida Spetnerin teoriaa: Esimerkiksi havainnot, joissa Spetnerin m��ritelm�n mukainen informaatio lis��ntyy ei falsifioi h�nen malliaan, koska ne voidaan tulkita "tuntemattomasta potentiaalista" johtuviksi. Spetner ei my�sk��n sovella evoluutioon soveltamaansa "Voisiko-se-toimia?" -kaavaansa omaan informaatiomalliinsa: l�hinn� koska h�nen mallinsa ei ainakaan toistaiseksi kerro mit��n sellaista mit� evoluutiokaan ei kertoisi - h�nen mallinsa keskeisimm�t, eli "mutaatioita ohjaava voima" ja "mikroevoluution potentiaali" kun ovat molemmat paitsi tuntemattomia my�s tuntemattoman suuruisia.
Kaikesta huolimatta Spetnerin malli ansaitsee erikoishuomion siit�, ett� se on toistaiseksi ainut kreationistien informaatiomalli, joka on sek� formalisaatio ett� vaihtoehtoinen malli.
Lis�tietoja:
Timo Tiainen,"Kriittinen arvio Spetnerin n�kemyksist�",2004
Ian Musgrave,"Information Theory and Creationism - Spetner and Biological Information",2005
Gert Korthof,"Could it work?",2003
Mark Chu-Carroll, "Using Bad Math to Create Bad Models to Produce Bad Results",2007
(Lee Spetner, "Not by Chance!", 1998, sivu 210)
Spetnerin keskittyy evoluutiokritiikkiin, mutta muista poiketen esitt�� my�s vastahypoteesin, "NREH":n.
*Evoluutiohypoteesissaan Spetner nojaa omaan informaatiom��rittelyyns�, joka ei ole kovin osuva -Spetner ei esimerkiksi k�sit�, ett� entsyymien ja substraattien sitoutumisella ei ole "kaikki tai ei mit��n" -luonnetta. H�n my�s laittaa yht�l�isyysmerkit "puhtaan Sattuman" ja "evoluution" v�liin. Spetnerin k�ytt�m� "hienos��t�perustelu", joka sallii informaation kasvun taas on ominaisuuksiltaan kovasti samanlainen, kuin luonnonvalinta.
*NREH:ssa keskeisint� on "evoluution ennalta ohjelmoitu potentiaali", joka m��r�� miten kehitys menee ja mink�laista informaatiota tulee. Spetner ei kuitenkaan selit� miten mutaatiot kaivavat potentiaalista informaatiota, eik� sit� miten t�t� potentiaalia voitaisiin tutkia. H�n paljastaa, ett� osa on "nukkuvia geenej�", mutta t�m� ei ole ainut menetelm�. Spetner ei my�sk��n kerro, syntyyk� "NREH":n piiriss� uutta informaatiota vai ei. Emme loppupeleiss� tied� mik� t�m� "potentiaali" on, emmek� my�sk��n mitk� ovat "NREH":n rajat, ja t�t� kautta kautta meill� ei ole keinoa falsifioida Spetnerin teoriaa: Esimerkiksi havainnot, joissa Spetnerin m��ritelm�n mukainen informaatio lis��ntyy ei falsifioi h�nen malliaan, koska ne voidaan tulkita "tuntemattomasta potentiaalista" johtuviksi. Spetner ei my�sk��n sovella evoluutioon soveltamaansa "Voisiko-se-toimia?" -kaavaansa omaan informaatiomalliinsa: l�hinn� koska h�nen mallinsa ei ainakaan toistaiseksi kerro mit��n sellaista mit� evoluutiokaan ei kertoisi - h�nen mallinsa keskeisimm�t, eli "mutaatioita ohjaava voima" ja "mikroevoluution potentiaali" kun ovat molemmat paitsi tuntemattomia my�s tuntemattoman suuruisia.
Kaikesta huolimatta Spetnerin malli ansaitsee erikoishuomion siit�, ett� se on toistaiseksi ainut kreationistien informaatiomalli, joka on sek� formalisaatio ett� vaihtoehtoinen malli.
Lis�tietoja:
Timo Tiainen,"Kriittinen arvio Spetnerin n�kemyksist�",2004
Ian Musgrave,"Information Theory and Creationism - Spetner and Biological Information",2005
Gert Korthof,"Could it work?",2003
Mark Chu-Carroll, "Using Bad Math to Create Bad Models to Produce Bad Results",2007
Lis�tietoja:
Timo Tiainen,"Kriittinen arvio Spetnerin n�kemyksist�",2004
Ian Musgrave,"Information Theory and Creationism - Spetner and Biological Information",2005
Gert Korthof,"Could it work?",2003
Mark Chu-Carroll, "Using Bad Math to Create Bad Models to Produce Bad Results",2007