6. Toπογραφική ανάλυση της εκφύλισης
στον πόρο του Müller
Αν χώρίσουμε με
νοητές γραμμές τον πόρο του Müller, διαπιστώνουμε τρία μέρη: το κεφαλικό μέρος το
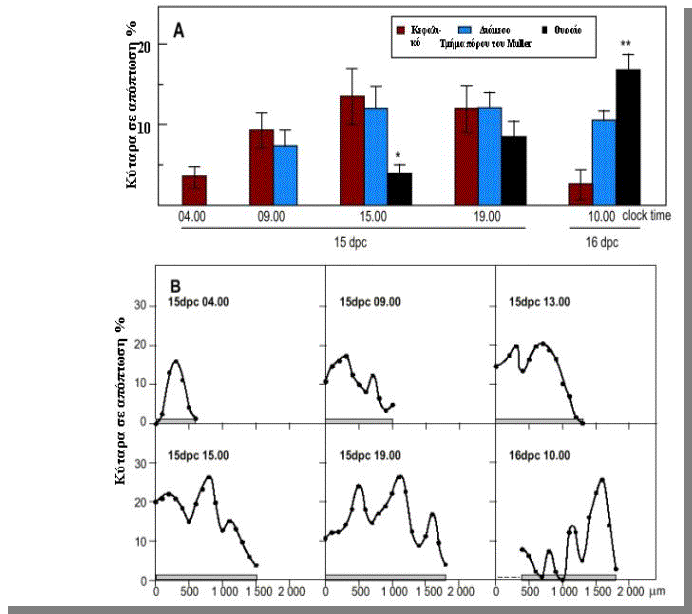
διάμεσο και το ουραίο μέρος. Στο σχήμα
5Α, φαίνεται παραστατικότατα η διαφορά στα επίπεδα της απόπτωσης των
κυττάρων στον πόρο του Müller. Aντιπροσωπευτικά προφίλ απόπτωσης
αποδεικνείουν ότι η απόπτωση λαμβάνει μέρος προοδευτικά κατά μήκος του πόρου του Müller, από το κεφαλικό
προς το ουραίο τμήμα του. Αυτό φαίνεται στο σχήμα 5Β στο οποίο η κορυφή
της καμπύλης αντιπροσωπεύει το μέγιστο ποσοστό επί τις εκατόν κυτάρων σε
απόπτωση κατά μήκος του επιμήκη άξονα των πόρων του Müller. Βλέπουμε προοδευτική μετακίνηση της
κορυφής της καμπύλης προς το ουραίο τμήμα.
H εμφάνηση του υποδοχέα της ΑΜΗ
συσχετίζεται με ένα κεφαλικό προς ουραίο τμήμα από απόπτωση τύπου Ι και με την
εκφύλιση του πόρου του Müller
Η ΑΜΗ επάγει
εμμέσως την εκφύλιση των πόρων του Müller διαμέσου του μεσεγχυματικού ιστου. Επιπρόσθετα η
ΑΜΗ δεν έχει επίδραση σε απομονωμένα Κύτταρα του πόρου του Müller ή δε πόρους οι
οποίοι καλλιεργόνται με ετερόλογο μεσέγχυμα το οποίο παρουσιάζει έλλειψη σε
υποδοχείς της ΑΜΗ. Τα mRNA της AMHRII εντοπίζονται στο μεσέγχυμα στην περιοχή γύρω από τους πόρους του Müller και σε θηλυκα και
σε αρσενικά έμβρυα αρουραίου την 14η ημέρα. Πρώτα εμφανίζονται στο
κεφαλικό τμήμα και ακολούθως επεκτείνονται προς το ουραίο καθως ο πόρος του Müller αυξάνεται προς το
ουραίο τμήμα. Η ανίχνευση με ειδικό αντίσωμα απέναντι στον ΑΜΗRII επιβεβαιώνει μια παρόμια κλίση συγκέντρωσης από
το κεφαλικό προς το ουραίο άκρο, το αποίο εμφανίζεται περίπου 10 ώρες μετά τα
μετάγραφα. Διαφορές κατά την κατανομή του μεσεγχυματικού ΑΜΗRII στο χώρο παρουσιάζονται μεταξυ αρσενικών και
θηλυκών εμβρύων. Στο αρσενικό αυτές οι μεταβολές οφείλονται λόγω στην ελάττωσης
τηε έκφρασης του AMHRII καθώς προχωρεί η διαδικασία της εκφύλισης
των πόρων του Müller. Η έκφραση του AMHRII απαιτεί άθικτο επιθήλιο και εξαρτάται από το σήμα
το οποίο λαμβάνεται από επιθηλιακά κύτταρα Wnt7a. Oι Parr και ΜcMahon (1998) έδειξαν ότι το αρσενικό
έβρυο ποντικιού το οποίο παρουσιάζει ανεπάρκεια στο Wnt7a, αποτυγχάνει να υποπέσει σε εκφύλιση των
πόρων του Müller λόγω αποτυχείας έκφρασης του υποδοχέα του ΑΜΗ. Η έλλειψη του Wnt7a λόγω της εκφύλισης
του επιθηλίου των πόρψν του Müller είναι υπεύθυνο για την προοδευτική μείωση του AMHRII στο μεσέγχυμα των πόρων του Müller κατά την
κεφαλουραία εκφύλισή των.
Χρησιμοποιώντας
την ευαίσθητη μέθοδο in situTUNEL, εντοπίστηκαν με ακρίβεια οι χωροχρονικές μεταβολές της αποπτωτικής
διαδικασίας και παρουσιάζονται περιληπτικά στο σχήμα 3, και επίσης
παρουσιάζεται το γεγονός του ότι η απόπτωση είναι μια προοδευτική διαδικασία
και όχι ταυτόχρονη. Έτσι ένα κύμα κυτταρικού θανάτου προχωράει από το κεφαλικό
προς το ουραίο άκρο, και σχετίζεται στατιστκά με την κεφαλουραία εμφάνιση των AMHRII.

H απόπτωση τύπου ΙΙ λόγω
διάρρηξης της βασικής μεμβράνης προωθεί την επιθηλιομεσεγχυματική μεταμόρφωση
Η εκφύλιση των
πόρων του Müller πυροδοτείται ή είναι στενά συνδεδεμένη με αλλαγές στο περιβάλλον μεσέγχυμα
και επιπρόσθετα, αλλαγές στην εξωκυτάρια ύλη στην επιθηλιο-μεσεγχυματική
επιφάνεια. Η βασική μεμβράνη η οποία περιέχει λαμινίνη και κολλαγόνο τύπου ΙV διατηρεί τη διαφοροποιημένη μορφολογία των
επιθηλιακών κυτάρων. Η αποδόμηση της λαμινίνης ή του κολλαγόνου τύπου IV πιθανόν να μπορεί να αρχίσει μια αποπτωτική
διαδικασία κυτταρικού θανάτου όπως έχει
περιγραφεί στο αμνίο αρουραίου (Lei et al.,1997, 1999) ή
στους γαλακτοφόρους αδένες. Έτσι η απώλεια επαφής με ορισμένα μόρια της εξωκυτταρίου
ύλης αναγνωρίζεται ως προαποπτωτικό σήμα (Frish and Francis, 1994; Meredith et al., 193).
Όταν τμήματα της βασικής μεμβράνης χαθούν και τα Κύτταρα γίνουν αποπτωτικά
τότε προκαλούνται αλλαγές στην οργάνωση του επιθηλίου. Ένας αριθμός επιθηλιακών
κυτάρων μετασχηματίζονται σε μεσεγχυματικά τα οποία είναι ικανά να
μεταναστεύσουν διαμέσου του συνδετικού ιστού. DiI-σημασμένα επιθηλιακά Κύτταρα των πόρων του Müller παρατηρήθηκαν να μεταναστεύον έξω από τον
πόρο σε αρσενικά έμβρυα αλιγάτορα. Η πρωτεύουσα αλλαγή στις πρωτεϊνες του κυτταροσκελετού
των νέων μεσεγχυματικών κυτάρων, είναι η απώλεια της κυτοκετατάνης η οποία
είναι απανταχού παρών στα επιθηλιακά Κύτταρα και η ταυτόχρονη εμφάνιση της βιμεντίνης η οποία χαρακτηρίζει
τα μεσεγχυματικά Κύτταρα. Έτσι η επιθηλιομεσεγχυματική μεταμόρφωση
χρησιμοποιείται για να εκμεταλλευτεί και να ανασχηματίσει ανεπιθύμητα
επιθηλιακά Κύτταρα, όπως περιγράφηκε και για τα έσω άκρα της υπερώας. Η
επιθηλιομεσεγχυματική μεταμόρφωση είναι ένα κύριο και πολυ σημαντικό γεγονός
για την εκφύλιση των πόρων του Müller.
Δράση και ρόλος της ΑΜΗ κατά την εκφύλιση
των πόρων του Müller
Θηλυκές ουρογεννητικές οδοί αρουραίου
καλλιεργήθηκαν με ανθρώπινη ανασυνδιασμένη AMH σε τελική συγκέντρωση 3μg/ml στη 14η μέρα για 24, 30 και 38
ώρες. Η απόπτωση που παρατηρήθηκε ήταν
όμοια και μπορούσε να συγκριθεί με τα αρσενικά έμβρυα στα οποία η ΑΜΗ είχε
παραχθει στους όρχεις. Παρατηρήθηκε έτσι με τη μέθοδο TUNEL θηλικοί και αρσενικοί ουρογενητικοί οδοί (Σχήμα 6Β,C)
Και τα ποσοστά των αποπτούμενων κυτάρων
στα θηληκα (16.2±2,7%)
μπορούσε να συγκριθεί με τα ποσοστά που λήφθησαν από αρσενικά έμβρυα (16.8±2,9%) όπως φαίνεται στο σχήμα 6Α. Τα
ποσοστά απόπτωσης στους πόρους του Wolff ήταν σημαντικά χαμηλότερα όπως φαίνεται στο σχήμα
4,Α.
Για την καλύτερη διαπίστωση του ρόλου που
παίζει η ΑΜΗ στους πόρους του Müller έγιναν πειράματα με διαγονιδιακά ποντίκια 14
ημερών, τα οποία παρουσίαζαν ανθρώπινο γονίδιο για την ΑΜΗ (MT-hAMH) και άλλα με
αφαιρεμένο το γονίδιο τα οποία δεν εξέφραζαν την ΑΜΗ (knockout) AMH-KO σε σύγκριση με
φυσιολογικά (wild type) ποντίκια. Τα ποσοστά απόπτωσης, όπως φαίνεται στο σχήμα 7,Α ήταν
σαφώς ψηλότερα στα αρσενικά έμβρυα. Αξιοπρόσεκτο είνιαι ότι παρουσιάστηκαν
μερικά ποσοστά απόπτωσης της τάξεως 2,8±0.6%. Σε αρσενικά και θηλυκά διαγονιδιακά ποντίκια τα
οποία υπερέκφραζαν την ΑΜΗ (ΜΤ-hAMH).
Παρουσιάστηκε ψηλότερο του κανονικού ποσοστο απόπτωσης που όπως φαίνεται
στο σχήμα φτάνει το 30%, και τα ποσοστά σχετίζονται (correlation) με τα ποσοστά ΑΜΗ στον ορό.
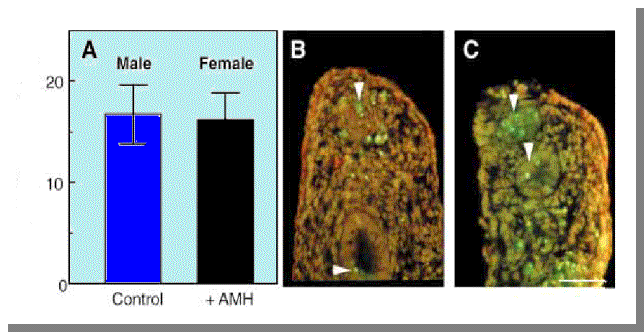
Σχήμα 6
A : Oυρογενητικά φύματα από 14 ημερών έμβρυα αρουραίων τα οποία καλλιεργήθηκαν για 18 ώρες. Όπως φαίνεται στη γραφική παράσταση παρουσιάζεται το % ποσοστό επιθηλιακών κυτάρων
στους πόρους του Müller τα οποία έχουν αποπέσει σε αρσενικό φυσιολογικό έμβρυο και σε Θηλυκό
του οποίου έχουν καλλιεργηθεί με ανασυνδιασμένη ΑΜΗ ανθρώπου
(hAMH).
B,C: Oυρογενητικές οδοί αρσενικου( B ) και Θηλυκού επεξεργασμένου με ΑΜΗ (C). Βλέπουμε καθαρά ότι παρατηρείται απόπτωση των πόρων του Müller και στα θηλυκά έμβρυα.
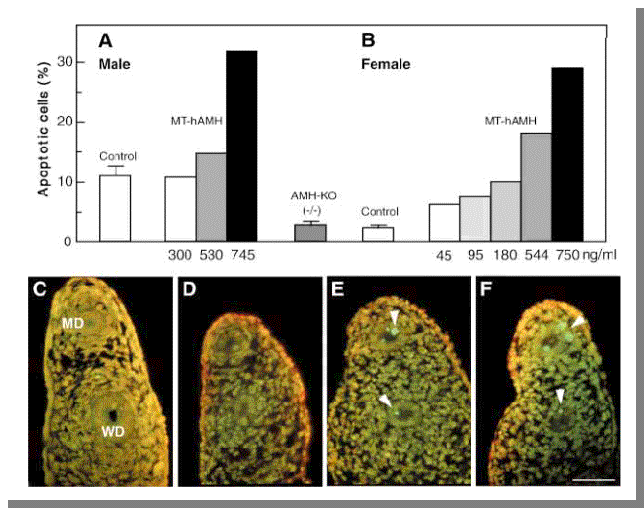
Σχήμα 7
A-B: Eπίδραση της ΑΜΗ in vivo σε πόρους του Müller σε άγριου τύπου(wild type) και διαγονιδιακά ποντίκια 14 ημερών. Στα διαγονιδιακά ποντίκια υπάρχουν
δίαφορα επίπεδα hAMH
που εκφράζονται σε ng/ml.Aξιοπρόσεκτο
είναι ότι τα ποντίκια με Knock Off (AMH-KO) προσεγγίζουν τα επίπεδα των θηλυκών Wild-type.
C-F: Σήμανση ΤUNEL. C-KnockOff D-Θηληκό E-Aρσενικό
με ΜΤ-hAMH 300ng/ml F-Θηλυκό με
MT-hAMH 180ng/ml.
Ο ρόλος β-Catenin (σχήμα 12) ως μεσολαβητή για την
εκφύλιση των πόρων του Müller που
οφείλεται στη δράση της ΑΜΗ
Η β-κατενίνη είναι μια
πολυλειτουργική πρωτεϊνη η οποία
δύναται να επηρρεάσει τόσο την κυτταρηή προσκόληση σε γειτονικά όσο και τη
μετάδοση σημάτων.(Resnik, 1997; Willert and Nusse, 1998). Ο διπλός αυτός ρόλος προυποθέτει τη
συνεργασία της με διαφορετικά
επικουρικά μόρια σε διαβορετικά υποκυτταρικά διαμερίσματα. Έτσι η β-catenin (σχήμα 12) μπορεί να βρεθεί τόσο στην πλασματική μεμβράνη,
όσο και στο κυτοσόλιο καθώς και στον πυρήνα. Ο ρόλος της στην κυτταρική προσκόλληση έχει καλά
καθιερωθεί. Η β-catenin, (σχήμα 12) συνεργάζεται σε
πεμπτοταγείς δομές πρωτεϊνών οι οποίες υπάγονται σε διαδικασίες κυτταρικής
προσκόλλησης (adhension), και ενώνοντας διαμεμβρανικές καντερίνες
(cadherines), με το εσωτερικό δίκτυο μικρονηματίων
ακτίνης. (Aberle et
al., 1996; Kemler, 1993). Αυτές
οι αλληλεπιδράσεις επιτρέπουν την κυτταρική προσκόλληση των κυτάρων μεταξύ τους
επάγει την πόλωση των κυτάρων και τη συγκρότηση των διακυτταρικών συνάψεων. H b-catenin (σχήμα 12) επίσης δρα
ως μεταγραφικός παράγοντας – ρυθμιστής και υπάρχουν ενδείξεις ότι μεσολαβεί στη
μετάδοση του σήματος Wnt (WiNgless-Type MMTV intergration site) (σχήμα 13).
Mετα το σήμα wnt, αρχίζει ένας καταρράκτης αντιδράσεων ο οποίος τελικά έχει σαν τελικό
αποτέλεσμα την αύξηση τηςκυτταροπλασματικής συγκέντρωσης της β-catenin. Επίσης με μηχανισμούς αρνητικής ανάδρασης, το
σήμα wnt(σχήμα 13) το οποίο υπενθιμίζεται ότι υποβοηθείται από τη β-catenin (σχήμα 12),
ενεργοποιεί την ρύθμιση της ποσότητας wnt που σχηματίζεται,
αφού διαμέσου της οδού ουμπικιτίνης και πρωτεασωμάτων, αποδομεί τη β-catenin. Μερά, υπάρχει μια επακολουθούμενη μετατόπιση της
β-catenin στον πηρύνα όπου εκεί αλληλεπιδρά με LEF (lemfoid enhancer factor) (σχήμα 14) / TCF (T-Cell Factor) παράγοντες, οι οποίοι ανήκοι στην οικογένεια των
μεταγραφικών παραγόντων οι οποίοι δημιουργούν ένα σύμπλοκο ικανό να
ενεργοποιήσει ή να αναστείλει τη μεταγραφή του γονιδίου. (Bedhers et al, 1996; Waltzer and
Bienz, 1999; Wodarz and Nusse, 1998). Το wnt είναι μόρια-σηματοδότες τους οποίους το
όνομα έχει ταυτιστεί με γεγονότα μορφογένεσης. Τα wnt1, wnt2, wnt3 και wnt3a, επάγουν
αποδεδειγμένα μια κυτταροπλασματική συγκέντρωση β-catenin. Tα υπόλοιπα wnt, τα οποία δεν ασκούν, ή ασκούν ελάχιστο
ρόλο στην κυτταρική μορφολογία δεν επηρεάζουν την συγκέντρωση της β-catenin. Το wnt4 για παράδειγμα
είναι
απαραίτητο για το σχηματισμό των πόρω
του Müller, ενώ το wnt7a, το οποίο εκφράζεται
από το επιθήλιο των πόρων του Müller, επιτρέπει τη διμορφική ανάπτυξη στα δύο φύλα,
αφού μετατρέπει με τέτοιο τρόπο το μεσέγχυμα, ώστε να είναι ικανό να αντιδρά στην ΑΜΗ μέσω του υποδοχεά της
ΑΜΗ.(Parr and McMahon, 1988).
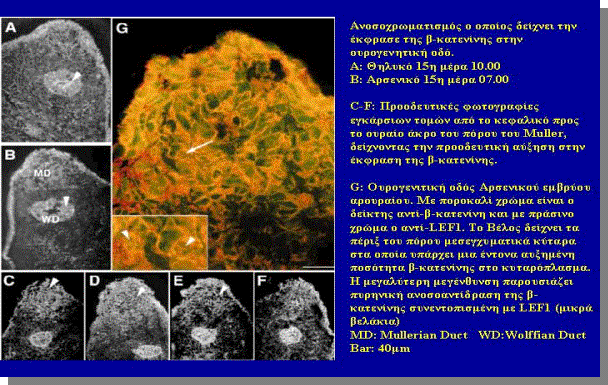
H AMH επάγει τόσο in vivo, όσο και in vitro, μια κυτταροπλασματική αύξηση της β-catenin (σχήμα 12) στα
μεσεγχυματικά Κύτταρα που υπάρχουν στην περιφέρεια του πότου του Müller (ίδε σχήμα 3).
In vivo, η παρουσία β-cateninin, παρατηρείται στα μεσεγχυματικά Κύτταρα των
αρσενικών ουρογεννητικών οδών κατά την περίοδο ευαισθησίας στην ΑΜΗ και
ολόκληρη η παρουσία της β-κατενίνης συσχετίζεται στατιστικά με την κεφαλουραία παρουσία των υποδοχέων AMHRII. Η κυτταροπλασματική β-κατενίνη πότε δεν
παρατηρείται σε φυσιολογικό θηλυκό έμβρυο in vivo παρά μόνο όταν επεξεργασθεί με
ανασυνδιασμένη β-κατενίνη.
Η έφραση
του ΑΜΗRII κατά
μήκος του πόρου του Müller στο
περί του πόρου μεσέγχυμα
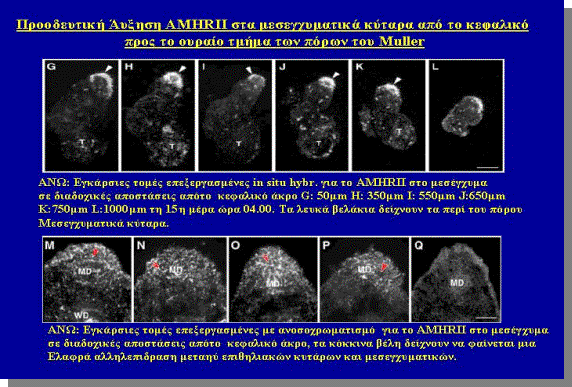
Mε μέθοδο της in situ υβριδοποίησης, κατά την οποία έγινε
εντόπιση του mRNA του AMHRII το οποίο εντοπίστηκε πρωταρχικά στα μεσεγχυματικά Κύτταρα γύρω από τον
πόρο του Müller στην 14η μέρα
ώρα 20.00 και τα επίπεδα του μετρήθηκαν με τη μέθοδο αρίθμησης της απεικόνησης
των ραδιενεργών κόκκων σε φωτογραφικο φίλμ από τη μέθοδο της in situ υβριδοποίησης με
ραδιενεργο 35S,
έμειναν σταθερά μέχρι την 15η μέρα ώρα 10.00 όπου αυξήθηκαν ελαφρά
(σχήμα 8,Α-F).
Κατά την αύξηση
του πόρου προς το ουραίο άκρο το AMHRII παρουσιάστηκε
στο γειτονικό μεσέγχυμα και πρώτα στο μεσέγχυμα κοντά στο καφαλικό άκρο και
ακολούθως στο μεσέγχυμα που βρίσκεται διαάμεσο και στο ουραίο άκρο (Σχήμα 8G-L). Αυτό αντανακλά και το προηγουμενο πείραμα (σχήμα 5,Β) κατά το οποίο
παρατειρείται προοδευτική απόπτωση επιθηλιακών κυτάρων, από το κεφαλικό προς το
ουραίο άκρο με στατιστικά σημαντική συσχέτιση.
Επίσης στα
σχήματα 8,Μ-Q ,
παρατηρείται ελαφρά αλληλεπίδραση μεσεγχυματικών κυτάρων και επιηθλίου
της επιφανείας Τελικά η σε σειρά προβολή των φωτογραφιών από κρυοστάτη μας
δείχνει τη συνολικά και γενικά παρατηρούμενη προοδευτική εμφάνηση της πρωτεϊνης
του υποδοχέα της ΑΜΗ τύπου ΙΙ από το κεφαλικό προς το ουραίο άκρο.
5.6.Ρόλος της
β-catenin (σχήμα 12) και του LEF1 (σχήμα 14) στην εκφύλιση των πόρων του Müller και το μοριακό μηχανισμό δράσης της ΑΜΗ
Για τη μελέτη της δράσης της β-catenin και του LEF1 (σχήμα 14),
έγινε ανοσοχρωματισμός σε διαδοχικές εγκάρσιες τομές της ουρογεννητικής
οδού σε έβρυα 14ων εώς 15
ημερών.
Και στα αρσενικά
και στα θηλυκά έμβρυα παρουσιάστηκε σε όλες τις χρονικές περιόδος μεγαλύτερη
εντόπιση β-catenin (σχήμα 12) στα επιθηλιακά
Κύτταρα της επιφάνειας των πόρων του Wolff παρά στους
πόρους του Müller. (Σχήμα 9,A-F) Κατά την 15η μέρα παρουσιάστηκε μια ισχυρή δέσμευση β-catenin στα μεσεγχυματικά Κύτταρα τα οποία
περιβάλλουν τους πόρους του Müller, και τα επιθηλιακά Κύτταρα των πόρων του Müller συνέχισαν να
έχουν στη μεμβράνη τους ενωμένη τη β-catenin. (Σχήμα 9,C-G)
Αξιοπρόσεκτο
είναι το γεγονός ότι παρατηρείται μόνο στα αρσενικά έμβρυα μια διαδοχική αύξηση
της β-catenin στα μεσεγχυματικά Κύτταρα από το κεφαλικό
προς το ουραίο άκρο των πόρων του Müller και όχι στα θηλυκά. (Σχήμα 9,C-F)
5.7.Επίδραση της ΑΜΗ στην έκφραση της β-κατενίνης στο μεσεγχυματικό στρώμα
Για τη διερεύνηση
του κατά πόσο η ΑΜΗ πυροδοτεί την έκφραση της β-κατενίνης (σχήμα 12) του
μεσεγχυματικού στρώματος, διεξάγει ανοσοχρωματισμός σε εγκάρσιες τομες
κρυοστάτη ουρογενητικών οδών αρσενικών και θηλυκών εμβρύων 14ων
ημερών τα οποία καλλιεργήθηκαν για 30 ώρες. Οι θηλυκές οθρογενητικές οδοί
καλλιεργήθηκαν με ή χωρίς παρουσιά ανασυνδιασμένη ανθρωπίνου αντιμυλερίου
ορμόνης σε τελική συγκέντρωση 3μg/ml. Παρουσιάστηκε κυτταροπλασματική συσσώρευση της
β-catenin (σχήμα 12) μόνο στα μεσεγχυματικά Κύτταρα θηλυκών εμβρύων τα
οποία είχαν επεξεργαστει με ΑΜΗ καθώς
και σε αρσενικά έμβρυα.
Αντίθετα σε
θυληκές ουρογεννητικές οδούς στις οποίες δεν υπήρχε ΑΜΗ δεν παρατηρήθηκε κυτταροπλασματική
συσσώρευση β-catenin .Έτσι παρουσιάζεται μια στατιστικά
σημαντική συσχέτιση μεταξύ παρουσίας ΑΜΗ και κυτταροπλασματικής εμφάνισης β-catenin (σχήμα 12). Eπίσης διπλός ανοσοφθορισμός έδειξε ότι στα
μεσεγχυματικά Κύτταρα που παρουσιάστηκε β-catenin παρουσιάστηκε και εντόπιση LEF1 (σχήμα 14) (lymphoid enhancer factor 1) στον πυρήνα
.
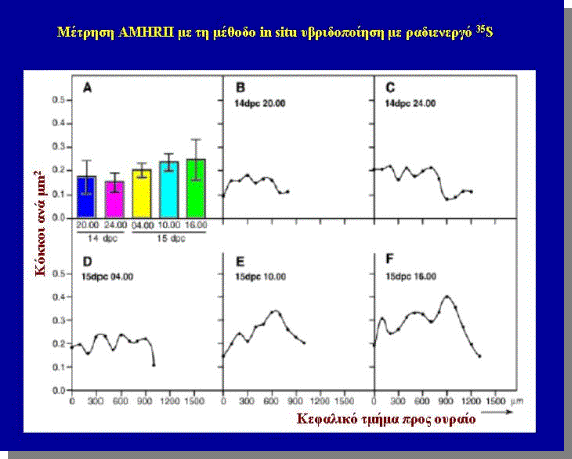
Σχήμα 8,A-F
Σχήμα 8,G-Q
Σχήμα 9. Παρουσία της β-κατενίνης στους πόρους του Müller.
Kυταροπλασματική και πυρηνική εντόπιση της
β-catenin (σχήμα 12)/επιβεβαιώθηκαν από πειράματα σε θηλυκά
μεσεγχυματικά Κύτταρα της ουρογεννητικής οδού. Αυτά τα Κύτταρα ληφθηκα με
μικροανατομή και αφού επεξεργάσθηκαν ανοσολογικλα ως προς τη βιμεντίνη και την
πρωτεϊνη ΑΜΗRII (σχήματα 1O,Α-Β), παρουσίασαν ισχυρή κυτταροπλασματική συσσώρευση
β-κατενίνης. Πολλά από αυτά τα κύτταρα παρουσίασαν β-κατενίνη και στον πυρηνα.
Αυτό το εύρημα μας κάνει να υποψιαζόμαστε την πιθανή δράση της β-κατενίνης στο
να μετεβάλλει τη μεταγραφή των γονιδίων στόχων και να
οδηγεί τελικά σε αλλαγές στην έκφραση των μεσεγχυματικών γονιδίων και τη μοίρα
των κυττάρων κατά την εκφύλιση των πόρων του Müller σε συνεργασία με το LEF1 (σχήμα 14) η οποία συνεντοπίστηκε σε πολλά απ’ αυτά τα κύτταρα μαζί με τη
β-κατενίνη (σχήμα 1O,C).
Πιέστε το πλήκτρο BACK στον πλοηγό σας για επιστροφή στο κυρίως
μενού.