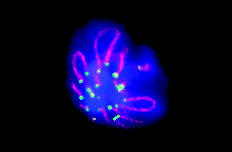
Telomeres (green) cluster for too long in spermatocytes lacking H2AX.
A histone variant involved in DNA repair and phosphorylated in response to critically short telomeres is nonetheless not involved in most aspects of telomere maintenance, according to Fernandez-Capetillo et al. (page 15). Their results suggest that this histone, H2AX, instead choreographs meiotic telomere movements.Telomeres at the ends of DNA strands are protected by proteins that prevent chromosome fusions. If these proteins are perturbed or if telomeres are severely shortened, the DNA damage response is triggered, repair factors are recruited to telomeres, H2AX is phosphorylated, and chromosome fusions and cell senescence often ensue. H2AX is a target of the ATM kinase, which itself helps to protect shortened telomeres from fusion events.
Given this background, the authors expected that H2AX might also function in telomere maintenance. Instead they found that H2AX mutant cells had telomeres of normal length and had normal chromosome fusion responses to shortened or deprotected telomeres. Telomeres were affected by H2AX loss, however, during a unique organizational stage in which telomeres cluster to form a bouquet-like structure that is associated with the onset of meiotic recombination.
Meiotic H2AX mutant cells were often unable to resolve the bouquet structure, and spermatocyte development frequently stalled at this stage. The group found that the H2AX phosphorylation that occurs in response to recombination- induced double-stranded DNA breaks is reduced in ATM mutants. Dephosphorylation of H2AX might signal that break repair has reached an advanced stage, recombination is near completion, and the bouquet structure can be resolved. This model would explain why development of ATM mutant spermatocytes also stalls in the bouquet stage. |