 Return to Biochemistry
Return to Biochemistry
CARBOHYDRATE METABOLISM
Download a copy of this study guide
 Return to top
Return to top
CARBOHYDRATE STRUCTURE
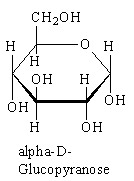
LACTOSE: beta-Galactose-1,4-Glucose.
- It is a disaccharide of glucose and galactose.
- It is considered a reducing sugar because it still has the free aldehyde group on C1 of
glucose.
- It hooks from the 1-carbon of galactose to the 4-carbon of glucose.
SUCROSE: alpha-Glucosyl-1,2-beta-Fructose
- A disaccharide of glucose and fructose.
Return to top
GLYCOLYSIS / FERMENTATION
RED BLOOD CELLS: They do only glycolysis. They have no mitochondria!
NET ENERGETICS OF GLYCOLYSIS: Net +2ATP come from glycolysis. 2 ATPs are invested
in the preparatory phase, and 4 ATPs (per original glucose) are recovered in the payoff phase.
METABOLISM AND DIFFERENT TISSUES: In nearly all tissues, glucose is trapped in the cell
by converting it to glucose-6-phosphate via the action of hexokinase or glucokinase.
- RED BLOOD CELLS
- Have no mitochondria. They continually produce lactate, which is continually
excreted.
- OH- Antiport: Lactate is continually excreted out of the RBC, in exchange for OH-
coming in. The net effect of this is to maintain the pH by preventing it from
becoming too acidic.
- BRAIN TISSUE
- Needs lots of glucose. It makes no lactate -- it has lots of mitochondria and it
metabolizes all of its glucose all the way to CO2.
- ADIPOSE TISSUE
- It can convert glucose-6-phosphate to glycogen
- It has mitochondria, but not very plentiful. Most of the glucose does not go through
TCA cycle but rather is used as building block for lipid biosynthesis.
- MUSCLE AND HEART TISSUE
- Can also make glycogen, but it cannot get back free glucose -- only glucose-6-phosphate.
- It will oxidize glucose fully as long as there is O2 around. If the muscle runs out of
oxygen, then fermentation will take over by mass action.
- Fermentation results in high lactate which can be excreted and which causes muscle
fatigue pain.
- LIVER PARENCHYMAL CELL
- Gluconeogenesis: The liver can use lactate, but first it turns it into pyruvate.
- Liver uses Glucokinase to trap glucose instead of hexokinase.
- Liver has a Glucose-6-Phosphatase that can actually restore free glucose, which it
then can secrete into the blood. The liver is the major source of blood-glucose
secretion.
- If the liver is making fat, then that is a sign of poor health.
GLUCOKINASE -VS- HEXOKINASE:
- Glucokinase has a higher km! It has a lower than normal affinity for trapping glucose in the
liver.
- This means it has a higher saturation point and a higher capacity for trapping glucose
via the kinase activity. This allows the liver to act as a buffer in taking up extra blood
glucose.
- Hexokinase is inhibited by its product, G-6-P, whereas glucokinase is not. This again allows
the liver to take up maximal amounts of glucose.
- Glucokinase is inhibited by F-6-P and F-1-P, whereas hexokinase is not.
CORI CYCLE:
- Muscle: Glucose ------> Lactate via glycolysis.
- Lactate goes to the liver, where it is taken up.
- Liver: Lactate ------> Glucose via gluconeogenesis
INVESTMENT PHASE OF GLYCOLYSIS: Steps 1 thru 3. It requires 2 molecules of ATP.
Overall: 1 Glucose ------> 2 Glyceraldehyde-3-Phosphate.
- Glucose + ATP ------> Glucose-6-Phosphate + ADP. Phosphorylation of Glucose.
- Hexokinase catalyzes the addition of phosphate from ATP to glucose. It works on
other hexose sugars too. It is an induced fit.
- Intermediate:
- Mg+2 is required as a cofactor in the substrate. The true phosphorylating
agent is the MgATP2- complex.
- Intermediate is an induced fit.
- First ATP comes from here.
- Glucose-6-Phosphate ------> Fructose-6-Phosphate.
- Reversible, catalyzed by phosphohexose isomerase.
- Changing a hexose to a ketose.
- Fructose-6-Phosphate + ATP ------> Fructose-1,6-biphosphate (FBP) + ADP
- Catalyzed by phosphofructokinase (PFK). This the primary regulatory enzyme.
- Inhibited when there is an excess of ATP.
- Second ATP comes from here.
- THIS IS IRREVERSIBLE -- THE COMMITTED STEP
FISSION STAGE: Stages 4 and 5 (or just 4, depending on how you'd like to divide it up).
- Fructose-1,6-biphosphate ------> Glyceraldehyde-3-Phosphate + Dihydroxyacetone
Phosphate -- Cleavage of Fructose-6-Phosphate.
- Catalyzed by Fructose Biphosphate Aldolase. It is a reversible aldol condensation.
- Dihydroxyacetone Phosphate <====> Glyceraldehyde-3-Phosphate
- So now we have made two glyceraldehyde phosphates. These guys are in reversible
equilibrium with each other, but the reaction is normally driven toward Glyc-3-Phosphate by mass action, i.e. that product is quickly taken up in the next step.
- The reversible equilibrium is catalyzed by triose phosphate isomerase.
THE PAYOFF PHASE: Steps 6 thru 10. Now that we have two molecules of Glyceraldehyde-3-Phosphate, all of the following occur twice per molecule of glucose.
- Glyceraldehyde-3-Phosphate + Pi + NAD+ ------> NADH + 1,3-bisphosphoglycerate --
Oxidation and phosphorylation.
- Catalyzed by Glyceraldehyde-3-Phosphate Dehydrogenase.
- An aldehyde is oxidized to a carboxylic acid.
- An inorganic phosphate group is attached.
- NAD+ ------> NADH -- It is a redox reaction, coupled to reduction of NAD+ to
NADH.
- The product is a an "acyl phosphate" -- a type of anhydride. It has very high
phosphorylation potential.
- 1,3-bisphosphoglycerate + ADP ------> 3-Phosphoglycerate + ATP
- Reaction is "coupled" with #6. Bisphosphoglycerate is the common intermediate.
- Produces *two* ATP per original glucose. This is the BREAK-EVEN POINT. We
have recovered our original investment.
- Very exergonic (DeltaG is even more negative than that required for ADP ------>
ATP). The product is quite stable.
- Remember that 1,3-bisphosphoglycerate is a powerful phosphorylating agent.
- Catalyzed by phosphoglycerate kinase.
- This is referred to as substrate-level phosphorylation, via a coupled reaction with
#6, as opposed to oxidative phosphorylation, via electron transport gradients.
- Or, it is just a direct transfer of phosphate from one compound to another.
- 3-Phosphoglycerate ------> 2-Phosphoglycerate. Simple isomerization.
- Catalyzed by phosphoglycerate mutase.
- Intermediate: 2,3-Bisphosphoglycerate (2,3-BPG).
- 2,3-BPG decreases hemoglobin affinity for oxygen.
- This step in the reaction can serve as a source for 2,3-BPG in the cell, in red
blood cells, e.g.
- When the phosphoglycerate mutase enzyme is bound to the membrane, it is
partaking in glycolysis, but when it is set free in the cytosol, it can give you
free 2,3-BPG to be used for other purposes.
- The enzyme can also function as a 2,3-BPG Phosphatase, working in reverse.
- 2-Phosphoglycerate ------> Phosphoenolpyruvate (PEP). Dehydration reaction.
- PEP has a high phosphorylation potential (much higher than reactant).
- Enzyme is enolase. Simple beta-removal of water.
- Phosphoenolpyruvate + ADP ------> Pyruvate + ATP
- The substrate is now completely dephosphorylated.
- 2 more ATPs produced per original molecule of glucose.
- Catalyzed by Pyruvate Kinase. Another substrate-level coupling reaction.
ENERGETICS OF GLYCOLYSIS: Total energy yielded per original molecule of glucose.
- +2 NADH: Glyceraldehyde-3-Phosphate Dehydrogenase (step 6)
- +2 ATP net
- -1 ATP (per original glucose): Hexokinase (step 1)
- -1 ATP (per original glucose): Phosphofructokinase (step 3)
- +2 ATP (per original glucose): Phosphoglycerate Kinase (step 7)
- +2 ATP (per original glucose): Pyruvate Kinase (step 10)
LACTIC ACID FERMENTATION: In the absence of oxygen, it allows for the NADH to be
reoxidized to NAD+. This is considered the official "end" to glycolysis, in the absence of oxygen.
- REDOX Reaction: One step process.
- REDUCTION: Pyruvate is reduced to lactate. (Keto changed to 2 alcohol).
- OXIDATION: NADH is oxidized to NAD+
- Note that overall process from Glucose to Lactate involves no net change in the oxidation
state of carbon. It is the same in glucose as in lactate.
- No ATP produced!!! The purpose is to regenerate the NAD+.
- Catalyst is lactate dehydrogenase (LDH). Note that although it is named dehydrogenase,
this reaction is actually a hydrogenation (hydrogens are added). Hence the enzyme is named
for the reverse reaction.
- This enzyme is used in REVERSE, too. Both reactions are essential:
- Pyruvate ------> Lactate: Gives us NAD+ for glycolysis and TCA cycle.
- Lactate ------> Pyruvate: Gives us NADH for electron transport chain, and
restores pyruvate for further metabolism.
- THE FATE OF LACTATE -- Ultimately the ONLY thing it can do is go back
to pyruvate! It can then be resynthesized back to glucose via
gluconeogenesis, or pyruvate can be oxidized via pyruvate dehydrogenase.
REGULATION OF GLYCOLYSIS:
- Metabolic Crossover: The notion that all intermediates before an inhibited step will tend to
build up, while all those intermediates after the step will tend to decrease.
- PASTEUR EFFECT: Glycolysis is substantially inhibited under aerobic conditions.
- The step that it is inhibited at is the Phosphofructokinase (PFK) step of Fructose-1-Phosphate ------> Fructose-1,6-Biphosphate
- O2 makes LDH work in reverse -- and fast -- to make pyruvate to be used in the TCA
cycle.
- NON-PHYSIOLOGICAL controllers:
- 2-Deoxyglucose -- No OH-group at 2'-position results in significant inhibition of
glucose.
- Sulfhydryl Reagents (Iodoacetic acid) -- they ruin glycerol phosphate dehydrogenase,
which has an essential sulfhydryl group.
- Fluoride Ion -- Potent inhibitor of enolase.
- The concentration required to inhibit glycolysis is orders of magnitude higher
than that found added in water or toothpaste.
- Arsenate -- phosphate analog will form 1-arsenate-3-phosphoglycerate which then
spontaneously hydrolyzes, to ruin glycolysis at the glyceraldehyde-3-phosphate
dehydrogenase step.
- REGULATORY ENZYMES:
- Hexokinase is inhibited by its product, Glucose-6-Phosphate.
- Phosphofructokinase (PFK) generally is inhibited by energy-rich indicators and is
stimulated by things that indicate a need for energy.
- Stimulated by: AMP, inorganic phosphate, Fructose-2,6-biphosphate
- Inhibited by: ATP, citrate, low pH (i.e. high levels of intracellular lactate).
- Fructose-2,6-biphosphate: A key regulating enzyme. Its levels are responsive to the effects
of insulin and glucagon.
- ACTION: It increase the rate of glycolysis.
- REGULATION: It is stimulated by insulin and inhibited by glucagon, which makes
sense if you think that insulin promotes the uptake and utilization of sugars.
FUTILE CYCLES -- generation of heat
- You can go back and forth between Glucose <====> Glucose-6-Phosphate, with the net
effect being the loss of ATP as heat. A different enzyme catalyzes it in each direction. This
is called a futile cycle.
- Malignant Hyperthermia -- an adverse reaction to Halothane Anesthesia in a small
percentage of people, leading to high fever and potentially death.
Return to top
OXIDATIVE DECARBOXYLATION / ELECTRON-TRANSPORT CHAIN
HIGH ENERGY COMPOUND STRUCTURE
- ATP
- Its anhydride bonds have a free energy of hydrolysis of 7.3 kcal / mol,. which is
therefore designated as the cutoff point for a high-energy compound.
- Almost always found as a magnesium salt. Mg+2 helps to neutralize the negative
charges of the phosphate.
- 3'-5' Cyclic AMP (cAMP)
- Has no high energy anhydride bond, but rather has a high-energy strained ring
structure.
- Phosphodiesterase cleaves cAMP ------> 5'-AMP. The equilibrium actually favors
5'-AMP.
- Caffeine inhibits phosphodiesterase, keeping cAMP levels high.
- ADP -- no natural reactions use ADP as an energy source, despite that it has a reactive
anhydride bond.
- ADP is utilized by converting it back to ATP, via the reaction: 2ADP ------> ATP
+ AMP
- 1,3-Bisphosphoglycerate
- Acetyl Phosphate
- Creatine Phosphate
- Found in skeletal muscle. It has a phosphoramide bond (P-O-N)
- Rapidly working muscles: Creatine Phosphate supplies depleted ADPs with a
phosphate group, to recharge them back to ATP.
- Creatine Phosphate is excreted in the urine at a constant rate proportional to muscle
mass. Measuring CP in the urine is a reliable indicator of muscle mass.
- Phosphoenol Pyruvate
- COENZYME-A: It has the following groups
- An ADP base.
- A Pantothenic Acid group, which must be supplied by the diet.
- A mercaptoethylamine group.
GLYCEROL-PHOSPHATE SHUTTLE: TRANSFERS NADH FROM CYTOSOL TO
MITOCHONDRIA.
- It results in production of FADH2 on the other side, which enters electron-transport chain in
the middle, thereby yielding only 2 ATP.
- Mechanism:
- OUTSIDE Membrane:
- Oxid: NADH ------> NAD+
- Red: Dihydroxy-Acetone Phosphate (DHAP) ------> Glycerol-3-Phosphate.
- Note this is NOT the same as the Glyceraldehyde-3-Phosphate in
glycolysis.
- The glycerol-3-phosphate then traverses the membrane.
- INSIDE Membrane:
- Oxid: Glycerol-3-Phosphate ------> DHAP
- Red: FAD ------> FADH2
MALATE SHUTTLE: TRANSFERS NADH FROM CYTOSOL TO MITOCHONDRIA
- It results in production of NADH on the other side, which enters at the beginning of the
electron-transport chain, thereby yielding 3 ATP.
- Mechanism:
- OUTSIDE Membrane
- Oxid: NADH ------> NAD+
- Red: Oxaloacetate ------> Malic Acid
- Malate then traverses the mitochondrial membrane.
- INSIDE Membrane
- Oxid: Malic Acid ------> Oxaloacetate
- Red: NAD+ ------> NADH
ELECTRON CARRIERS -- STRUCTURE
- NADH: Nicotinamide Adenine Dinucleotide
- Has a Nicotinamide Ring that carries the active carbon that is alternatively oxidized
and reduced.
- The ring is AROMATIC when oxidized and is NON-AROMATIC (in a
cyclohexodiene form) when reduced.
- We derive the nicotinic acid from our diet, from niacin.
- FADH2: Flavin Adenine Dinucleotide
- Derived from vitamin riboflavin
- Has a unique 3-ring isoalloxazine structure, in which the two active hydrogens are
found.
- Contains a ribitol alcohol group.
- FMN is similar to it.
THE ELECTRON TRANSPORT CHAIN: It occurs on the outer surface of the inner mitochondrial
membrane, through a series of membrane-bound electron-shuttles that pass electrons from one
complex to the next without letting go of intermediates.
- Enzymes involved:
- Flavoproteins
- Often contain Non-Heme iron, which interchanges between Fe+2 and Fe+3 forms.
- STEPS: Successive oxidations and reduction are approx. as follows:
| OXIDATION |
REDUCTION |
DESCRIPTION |
| NADH ------> NAD+ |
FMNH2 ------> FMN |
This first step is a two
electron transfer |
| FMN ------> FMNH2 |
Fe+3 ------> Fe+2 |
From here forward, only one
electron at a time is
transferred |
| Fe+2 ------> Fe+3 |
Quinone ------>
Hydroquinone |
Coenzyme-Q contains the
quinone group. THIS IS THE
MIDDLE STEP, WHERE
FADH2 IS ADDED IF
PRESENT |
| Hydroquinone ------>
Quinone |
Fe+3 ------> Fe+2 |
Cytochrome contains the Fe
this time, and various
cytochromes contain heme-
irons for the next several
steps. |
CYTOCHROMES: They are heme-containing proteins similar to hemoglobin. Different
cytochromes carry the electrons for the next several steps.
- Each time exchanging Fe(II) and Fe(III), we have: Cytochrome b-566 ------> Cytochrome
b-562 ------> Non-heme FeS protein ------> Cytochrome C1 ------> Cytochrome c
------> Cytochrome a a3.
- Cytochrome-a a3 is what finally reacts with oxygen to yield H2O
ATP SYNTHESIS AND PROTON PUMPING:
- Protons are pumped from the inside of the mitochondrial matrix outside, to the
intermembrane space.
- They then flow back into the matrix by an electrochemical gradient, and in so doing they
catalyze the synthesis of ATP by an ATPase working in reverse.
- The ATPase is integral to the inner mitochondrial membrane.
- pH Gradient: The electrochemical gradient can also be looked at as a pH gradient, where
the matrix is more basic and the intermembrane space is more acidic.
SUPEROXIDE ANION: O2-, a harmful partially reduced form of O2 (i.e. a peroxide derivative).
This is very reactive and harmful.
- We break it down as a defense mechanism, by a two-step process. This occurs in the
organelles, peroxisomes.
- Superoxide Dismutase: O2- ------> H2O2, aka hydrogen peroxide
- Catalase: H2O2 ------> H2O + O2
- CLINICAL -- newborns are sensitive to high oxygen tension, because they have not yet
developed superoxide dismutase.
- The retinal tissue is what is affected. Blindness may result.
- CLINICAL -- these oxidative processes are thought to play an important role in aging.
UNCOUPLERS: Agents that decrease the efficiency of the electron-transport ATP-synthase
gradient, causing a lower production of ATP and a greater amount of heat given off.
- Keep in mind that the production of ATP in the electron transport chain is not
stoichiometric. There is no 1:1 relationship, and the yields are approximate amounts based
on energetic calculations.
- Dinitrophenol (DNP): A well-known uncoupling agent. It would make a good diet pill as
an uncoupling agent (consume energy and give it off as heat rather than make ATP), except
it has terrible side effects -- severe cataracts.
- Thyroxine = the body's natural uncoupling agent. Thyroxine is released in response to cold
temperature to raise body temperature by acting as an uncoupler in the electron-transport
chain.
Return to top
PYRUVATE DEHYDROGENASE + THE TRICARBOXYLIC ACID CYCLE
OVERALL REACTION: The reaction occurs in the mitochondria.
- Pyruvate ------> Acetyl-CoA + CO2
- Multi-enzyme complex, the intermediates remain bound to the enzyme. Enzyme channeling.
- Overall reaction is irreversible, a REDOX Reaction.
- OXID: Pyruvate ------> Acetyl-CoA
- RED: NAD+ ------> NADH
COFACTORS REQUIRED FOR PYRUVATE DEHYDROGENASE: Five different coenzymes are
in the reaction:
- Thiamine pyrophosphate (TPP)
- From vitamin Thiamin
- Same as that which catalyzes Decarboxylation of Pyruvate ------> Lactic Acid in
Fermentation.
- It has an unusual 5-membered heterocyclic ring, containing a sulfur.
- Flavine Adenine Dinucleotide (FAD)
- From vitamin Riboflavin
- Coenzyme-A (CoA)
- From vitamin pantothenate
- Nicotinamide Adenine Dinucleotide (NAD+)
- From vitamin niacin
- Lipoate: Has two thiol (SH) groups.
- Oxidation of Lipoate yields two disulfide (S-S) bonds.
- Reduced Form: (SH) Lipoate acts as an acyl carrier.
- Oxidized form: (SS) Lipoate acts as an electron carrier.
- Lipoate is usually found bound to a Lysine of the enzyme it is a cofactor for.
Coenzyme-A: Has a reactive Thiol (SH) group.
- When acyl groups get hooked on (i.e. acetyl), they form a thioester with very high free
energies of hydrolysis.
PYRUVATE DEHYDROGENASE COMPLEX (PDC): The enzyme complex that catalyzes
conversion of pyruvate to Acetyl-CoA.
- (E1) PYRUVATE DEHYDROGENASE:
- COFACTOR: TPP (Thiamine Pyrophosphate)-- Thiazolium ring. It acts as an
electron sink -- it can take electrons into its ring and stabilize them.
- (E2) DIHYDROLIPOYL TRANSACETYLASE:
- COFACTORS: Lipoate, CoA
- The Core: E2 sits between E1 and E3.
- Lipoate: Its long chain hooks to a Lys residue on the enzyme.
- It has oxidized (disulfide) and reduced (-SH, -SH) forms, and can act both as
an acyl carrier and an electron carrier.
- Lipolysyl Groups: The name of the long chains at the end of the lipoate,
which are hooked to Lys residues on the enzyme.
- (E3) DIHYDROLIPOYL DEHYDROGENASE:
- COFACTORS: FAD, NAD
- FAD: The reactive part are the two CN bonds in the center of the ring.
OXIDATIVE DECARBOXYLATION (Pyruvate Dehydrogenase): FIVE STEPS, occurs
through substrate channeling. The enzyme complex never leaves the substrate!
- Step 1: Pyruvate (3C) decarboxylates to 2 Carbons
- Enzyme: Pyruvate Dehydrogenase (E1)
- CO2 comes off -- this is the FIRST CO2 that comes off from the original glucose.
- The remaining C=O group turn into a CH2OH group, and it hooks to TPP.
- Step 2: 2-Carbon-Alcohol oxidizes to carboxylic acid (Acetate)
- OXID: Alcohol ------> Carboxylic Acid
- RED: Lipoyl disulfide (-S-S-) group is reduced to 2 Thiols (2 -SH).
- Step 3: Esterification of the Acid with the Lipoyl group.
- Step 3a: Acetate is esterified to one of the Lipoyl -SH groups (to form SCH3CO2).
- Step 3b: The Ester from (a) then transesterifies with CoA-SH (The CoA has an SH
on it!) to form Acetyl-CoA, which is given off.
- Enzyme: Dihydrolipoyl Transacetylase -- This is the enzyme that yields Acetyl-CoA.
- Step 4: Lipoyl group is reoxidized
- OXID: Lipoyl-SH ------> Lipoyl-Disulfide (-S-S-)
- RED: FAD ------> FADH2
- Enzyme: Dihydrolipoyl Dehydrogenase
- Step 5: FAD is regenerated (reoxidized)
- OXID: FADH2 ------> FAD
- RED: NAD ------> NADH
REGULATION OF PYRUVATE DEHYDROGENASE:
- Pyruvate Dehydrogenase Kinase (PDH-Kinase) uses ATP to phosphorylate Pyruvate
Dehydrogenase to inactive it. So, the activity of PDH is controlled by phosphorylation.
- PHOSPHORYLATED FORM: Phosphopyruvate. INACTIVE
- Pyruvate Dehydrogenase Phosphatase (PDH-Phosphatase) dephosphorylates it.
- DEPHOSPHORYLATED FORM: Pyruvate Dehydrogenase. ACTIVE.
- The phosphatase is active all the time, so the ultimate phosphorylation control then
depends on the levels of the PDH-Kinase present.
- POSITIVE CONTROLS on the kinase: These things turn off production of PDH. They are
the products of PDH itself -- Acetyl-CoA and NADH.
- Increase Kinase Activity ------> Decrease PDH activity ------> Less Acetyl-CoA
is made.
- NEGATIVE CONTROLS on the kinase: These things turn on production of PDH. They
are the substrates of the PDH reaction -- Pyruvate, NAD+, and Coenzyme-A.
- Inhibit the kinase ------> Maintain PDH activity ------> More Acetyl-CoA is
made.
OVERALL ENERGETICS OF THE TCA CYCLE: You get an energetic equivalent of 12 ATP per
turn (i.e. 24ATP per original glucose) of the cycle, and 36 ATP total when you include Glycolysis.
| Item |
ATP Value |
Enzymatic Reaction |
| 2 NADH |
6 ATP |
Glyceraldehyde-3-Phosphate Dehydrogenase
(glycolysis) |
| 1 NADH |
3 ATP |
Isocitrate Dehydrogenase (TCA: form alpha-Ketoglutarate) |
| 1 NADH |
3 ATP |
alpha-Ketoglutarate Dehydrogenase (TCA: form
Succinyl-CoA) |
| 1 NADH |
3 ATP |
L-Malate Dehydrogenase (TCA: form Oxaloacetate) |
| 1 FADH2 |
2 ATP |
Succinate Dehydrogenase (TCA: form Fumarate) |
| 1 GTP |
1 ATP |
Succinate Thiokinase (TCA: form Succinate) |
| TOTAL: |
6 ATP per glucose from
glycolysis.
12 ATP per turn of
TCA Cycle, or 24 ATP
per glucose from TCA
cycle. |
|
| GRAND
TOTAL: |
36 ATP per original
glucose |
|
ACETYL-COA:
- Acetyl-CoA cannot lead to sugar.
- Sources of Acetyl-CoA
- Glucose (via pyruvate)
- Fats (via beta-Oxidation)
- Proteins
- Fates of Acetyl-CoA
- TCA Cycle
- Fatty Acid / Sterol Synthesis
- Ketone Bodies
STEPS OF THE TRICARBOXYLIC ACID CYCLE
- STEP I: Acetyl-CoA + Oxaloacetate ------> Citrate + CoEnzyme-A -- THE
COMMITTED STEP
- (2C + 4C ------> 6C)
- Condensation reaction.
- ENZYME: Citrate Synthase.
- INTERMEDIATE: Citroyl-CoA, which does not detach from enzyme. Then, CoA
comes off to yield Citrate + CoA.
- CoA has the thioester linkage with high free-energy of hydrolysis. This makes the
forward reaction favored and drives essentially the whole process.
- CoA is then recycled back to the Oxidative Decarboxylation system.
- Equilibrium in this reaction very much favors citrate.
- STEP II: Citrate ------> Isocitrate
- ENZYME: Aconitase.
- It contains an iron-sulfur center (prosthetic group)
- INTERMEDIATE: cis-Aconitate (2 3C intermediates). Does not dissociate from
enzyme.
- H2O can be added to cis-aconitate in two ways. One leads to citrate and the other
leads to isocitrate.
- Equilibrium reaction is driven to right by quick removal of isocitrate in next reaction.
- STEP III: Isocitrate ------> alpha-Ketoglutarate + CO2
- Oxidative Decarboxylation. Now we have a 5C compound.
- ENZYME: Isocitrate Dehydrogenase.
- Concurrent Reduction: NAD+ ------> NADH
- Intermediate: Oxalosuccinate -- not a free intermediate, but an enzyme-bound
chemical intermediate.
- STEP IV: alpha-Ketoglutarate ------> Succinyl-CoA + CO2
- Oxidative Decarboxylation. Now we have a 4C compound.
- ENZYME: alpha-Ketoglutarate-Dehydrogenase Complex.
- The reaction is similar to pyruvate dehydrogenase complex.
- Complex has three enzymes, analogous to E1, E2, and E3.
- OXID: alpha-Ketoglutarate ------> Succinyl-CoA
- Energy in conserved in the COS- (thioester) bond of -CoA.
- RED: NAD+ ------> NADH
- COFACTOR: Coenzyme-A, of course.
- Product is a thiol-ester -- a high-energy compound.
- STEP V: Succinyl-CoA ------> Succinate
- Cleavage of -CoA off of the Succinyl group, breaking a high-energy bond. This high-energy bond is coupled to formation of GTP.
- Substrate-Level Phosphorylation: Direct phosphorylation of a GDP to produce
GTP.
- GTP comes off here, instead of ATP!
- Note that this is the only time we get an ATP or GTP out of the TCA cycle.
All other times we get reduced cofactors (i.e. NADH)
- ENZYME: Succinate Thiokinase.
- LABEL-RANDOMIZING STEP: At this point we have a symmetrical compound.
Chirality is lost, and the two original carbons we invested as Acetyl-CoA will
therefore not be the same ones we get out.
- STEP VI: Succinate ------> Fumarate
- (Still 4C): Form a double-bond between the middle-two carbons. Fumarate is a
trans-dicarboxylic acid.
- ENZYME: Succinate Dehydrogenase.
- RED: FAD ------> FADH2
- INHIBITOR -- Malonate is a strong competitive inhibitor of the enzyme. It blocks
the citric acid cycle. It is the cis-acid of fumarate.
- STEP VII: Fumarate ------> L-Malate.
- Hydration Reaction: Add H and OH across the double bond.
- STEREOSPECIFIC: Only l-malate is formed, and only l-fumarate is a substrate.
- ENZYME: Fumarase.
- STEP VIII: L-Malate ------> Oxaloacetate.
- Oxidation: The -OH group is oxidized to a keto-group.
- ENZYME: l-Malate Dehydrogenase.
- Concurrent Reduction: NAD+ ------> NADH
REGULATION OF THE TCA CYCLE:
- Pyruvate Dehydrogenase: Inhibited by its products -- Acetyl-CoA and NADH.
- Part of the NADH influence is substrate deprivation -- that implies that you have low
levels of NAD+ which is required for the reaction.
- alpha-Ketoglutarate Dehydrogenase (alpha-Ketoglutarate ------> Succinyl CoA) -- this
is the key regulatory step.
- Ca+ stimulates the step, i.e. Ca+ promotes the TCA cycle.
- NADH and Succinyl CoA both inhibit the step -- two products of the reaction.
- High levels of GTP inhibit the alpha-Ketoglutarate Dehydrogenase Step.
ANAPLEROTIC REACTIONS: A reaction that provides an essential intermediate in central
metabolism.
- For example, in order for TCA cycle to function, we must have Oxaloacetate and Acetyl-CoA. Any source of these intermediates would then serve as an anaplerotic reaction.
- Alanine ------> Pyruvate is anaplerotic, because it provides us with pyruvate, a viable
starting point for TCA cycle.
- Pyruvate ------> Oxaloacetate (in Gluconeogenesis) -- this is anaplerotic, as it provides
us with essential Oxaloacetate.
Return to top
GLYCOGENESIS / GLYCOGENOLYSIS
STRUCTURE, FUNCTION, AND FLUCTUATION OF GLYCOGEN:
- Glycogen as an energy source:
- Fluctuating Levels: Glycogen levels increase and decrease markedly every day.
Generally they increase after a large meal, and decrease as we expend energy.
Glycogen levels are fairly depleted when we wake up in the morning.
- Glycogen is a SHORT-TERM energy source. Fat is the long-term energy source.
- GLYCOGEN STRUCTURE: Glycogen is a highly branched polymer of pure glucose.
- Linear Links: alpha-1,4 polyglucose links.
- Branched Links: alpha-1,6 polyglucose links
- Reducing Sugar: In a single polymer of glycogen, there is one sugar that still has a
reducing carbon on it at the 1 position. It is the "seed" of the polymer.
- BRANCHES -- approximately every fourth residue has a branch.
GLYCOGENOLYSIS: Breakdown of Glycogen to Glucose-1-Phosphate.
- Glycogen Phosphorylase: Glycogenn ------> Glucose-1-Phosphate + Glycogenn-1
- This is a Phosphorolysis reaction -- not hydrolysis!
- No ATP is required to break it down. But ATP was required to build it up, so we're
not getting something for nothing.
- Glycogen Phosphorylase cleaves only the alpha-1,4 linkages -- not the alpha-1,6
linkages.
- Glycogen phosphorylase takes any glucose chain down to the last four residues before a
branch point. It does not cut beyond that!
- Debranching Enzyme: Takes over when within four residues to a branch point. It has two
activities.
- 4-alpha-D-Glucanotransferase: Transfer 3 residues to another chain on the
polymer.
- Amylo-alpha-1,6-Glucosidase: Hydrolysis of the final 1,6 bond.
- This step creates FREE GLUCOSE -- not Glucose-1-Phosphate.
- This is a true hydrolysis reaction -- not phosphorolysis.
- Phosphoglucomutase: Glucose-1-Phosphate ------> Glucose-6-Phosphate.
- Isomerizes Glucose to a form that can be utilized in further metabolism.
- LIVER ONLY: Glucose-6-Phosphatase: Glucose-6-Phosphate ------> Glucose
------> Bloodstream.
- Modulation of blood-glucose level by the liver.
- Glucagon promotes higher blood glucose, and insulin promotes lower.
REGULATION OF GLYCOGENOLYSIS: Phosphorylation turns on the system.
- TWO FORMS OF GLYCOGEN PHOSPHORYLASE: ACTIVATE AND INACTIVE
- PHOSPHORYLASE-A: The activated glucose phosphorylase enzyme. It is
phosphorylated to be activated.
- Glucose and ATP have inhibitory effects on Phosphorylase-A. These guys
indicate excess energy, and we don't need to do glycogenolysis in that case.
- PHOSPHORYLASE-B: The inactive form of the glucose-phosphorylase enzyme.
It is dephosphorylated to be inactivated.
- AMP has a stimulatory effect on Phosphorylase-B, and can greatly increase
its activity, even without phosphorylation. AMP has no effect on
phosphorylase-a because its effect is overriden by the phosphorylation.
- TWO PHOSPHORYLATING ENZYMES
- PHOSPHORYLASE KINASE A (ACTIVATOR): A REGULATORY ENZYME
which uses ATP to phosphorylate Phosphorylase-B, in order to activate it. So, it has
an activating activity.
- cAMP activates Phosphorylase Kinase A, via a cAMP-dependent protein
kinase.
- Ca+2 (from IP3 transduction pathway) increases the activity of phosphorylase-kinase, as the delta-subunit of the enzyme is a calmodulin which, in this case,
increases the activity of the enzyme when bound to calcium.
- PHOSPHOPROTEIN PHOSPHATASE (DEACTIVATOR): ANOTHER
REGULATORY ENZYME which dephosphorylates Phosphorylase-A, in order to
inactivate it. So, it has an inhibitory activity.
- INHIBITOR-1A: Inhibits Phosphoprotein Phosphatase, in order to
promote (or sustain) glycogenolysis.
- Epinephrine and Glucagon cause the synthesis of 2nd messenger cAMP
------> caMP-dependent Protein Kinase ------> activates Inhibitor-1A
------> Inhibits Phosphoprotein Phosphhatase ------> PKA remains
phosphorylated ------> glycogen phosphorylase remains active.
- THE IMPORTANCE OF CAMP: It not only stimulates the conversion of phosphorylase-B
to phosphorylase-A, but it also inhibits the conversion of A to B.
- cAMP turns on Inhibitor 1A in order to inhibit the Phosphatase ------> Turn on
glycogen phosphorylase.
- Phosphorylates PKA directly ------> generate more Phosphorylase-A (Active
glycogen phosphorylase).
GLYCOGENESIS:
- YOU MUST START WITH ACTIVATED UDP-GLUCOSE
- Hexokinase / Glucokinase: Glucose ------> Glucose-6-Phosphate
- ATP required. Glucokinase used in liver, and hexokinase in muscle.
- Phosphoglucomutase: Glucose-6-Phosphate ------> Glucose-1-Phosphate
- This is the opposite reaction as what occurs at the end of glycogenolysis.
- Glucose-1-Phosphate Uridyltransferase: Glucose-1-Phosphate ------> UDP-Glucose
- UTP ------> UMP + PPi. When UMP hooks onto Glucose-1-Phosphate, it
then becomes UDP-Glucose.
- That means that UDP-Glucose has the UDP attached to Carbon #1!
- Pyrophosphatase: Cleaves the high-energy phosphoric anhydride bond.
This drives the formation of UDP-Glucose and makes its formation
irreversible.
- GLYCOGEN SYNTHASE: Glycogenn + UDP-Glucose ------> Glycogenn+1 + UDP
- Glycogen synthase requires a primer! It will add another glucose residue to an
already-existing chain.
- Glycogen synthase makes only linear glucose, connected in alpha-1,4 linkages. A
branching enzyme is required for branching.
- BRANCHING ENZYME:
- Once the glycogen chain is about 11 residues long, the branching enzyme removes
about 7 of the residues.
- It places the 7-residue chain in a 1,6-linkage, at least 4 residues away from any other
branching point.
- GLYCOGENIN: The self-glycosylating protein that serves as the primer for de novo
glycogen synthesis.
- Usually, no primer is needed:
- Glycogen Synthase normally has a low Km (i.e. highly sensitive), given a large
starting glycogen molecule.
- The larger the starting substrate, the lower the Km for Glycogen Synthase.
It won't start with glucose alone because the Km is way-the-hell too high.
- GLYCOGENIN ACTIVITY: It has a Serine OH-Group onto which it adds the first
glucose, to start the ball rolling.
- It takes the first glucose from activated UDP-Glucose, releasing UDP (just
like the synthase).
- Glycogenin then dissociates from the growing glycogen molecule after a few
residues have been put on.
REGULATION OF GLYCOGENESIS: Phosphorylation turns off the system.
- TWO FORMS OF THE GLYCOGEN SYNTHASE ENZYME: ACTIVE AND INACTIVE.
- GLYCOGEN-SYNTHASE A: ACTIVE FORM. It is dephosphorylated.
- GLYCOGEN-SYNTHASE B: INACTIVE FORM. It is phosphorylated.
- Glucose-6-Phosphate stimulates Glycogen-synthase-B to some extent. A
surplus of G-6-P indicates that glycogenesis should take place.
- INHIBITORY EFFECTS / PHOSPHORYLATION: Epinephrine and/or Glucagon are
secreted at times of low blood sugar or when we need cellular energy. They cause a
phosphorylation cascade.
- cAMP, DAG, and Ca+2 (i.e. beta-adrenergic secondary messengers) all shut off
glycogen synthase by promoting the phosphorylated form of the enzyme. There are
two mechanisms:
- Direct phosphorylation of glycogen synthase itself, thereby inactivating it.
- Phosphorylation of Inhibitor-1 a ------> phosphorylate (inhibit) the
phosphatase ------> glycogen synthase remains phosphorylated and hence
inactive.
- STIMULATORY EFFECTS / DEPHOSPHORYLATION: Glucose stimulates the
production of glycogen.
- Glucose inhibits Glycogen Phosphorylase-A and the phosphoprotein phosphatase,
as noted above.
- Phosphorylase-A inactivates Glycogen Synthase. It is therefore necessary
to inactivate the phosphorylase before we can make glycogen.
- Glucose therefore stimulates glycogen synthesis by inhibiting
glycogenolysis.
- Glucose also stimulates Phosphoprotein Phosphatase.
- Phosphoprotein Phosphatase itself promotes glycogen synthesis. It
promotes the dephosphorylated (active) form of Glycogen Synthase.
INSULIN:
- It acts by an autophosphorylating tyrosine-kinase receptor protein.
- It is generally anabolic: it promotes glycogen synthesis and inhibits glycogenolysis.
- It is secreted when glucose levels in the blood are high, and it promotes uptake of glucose by
most cells.
- It does not affect glucose-uptake by liver cells, because the liver can uptake as much glucose
as it wants by facilitated transport. Liver absorption of glucose is independent of insulin
levels.
OVERALL ENERGETICS OF GLYCOGEN:
- GLYCOGENOLYSIS: Every glucose residue released gives us +3 ATP. The end-product
of glycogenesis is glucose-6-P, which can then go through glycolysis.
- Because the product is glucose-6-phosphate, we don't need hexokinase in the first
step of glycolysis so we save ourselves 1 extra ATP, giving us a net of +3 ATP
(rather than +2) once the glucose has undergone glycolysis.
- GLYCOGENESIS: It costs -2 ATP to add on one residue of Glucose to a glycogen chain.
It costs ATP to make glycogen.
- 1 ATP cost from glucokinase.
- 1 ATP cost (via UTP) for the glucose-1-phosphate uridyltransferase
- NET ENERGY YIELD: ONLY +1 ATP PER GLUCOSE FROM GLYCOGEN.
- WHY MAKE GLYCOGEN?
- We make glycogen in times of plentiful energy. We break it back down when we
need quick energy. Gynogen can be broken down quickly because it is highly
branched.
- We actually get more ATP (3 -vs- 2) from glycogen then we do from free glucose, if
we disregard the ATP originally invested.
GLYCOGEN STORAGE DISEASES:
- Type-I (Von Gierke's Disease): Deficiency of Glucose-6-Phosphatase in liver, intestinal
mucosa, and kidney
- Symptoms:
- Hypoglycemia -- liver can't release glucose!
- Lactic Acidemia -- Inability to process lactose normally, as well as increased
glycolysis.
- Increased Uric Acid -- cause not clear.
- Treatment: Taking small amounts of carbohydrate during the day to maintain blood
glucose can help diminish symptoms.
- Type-II (Pompe's Disease): Accumulation of glycogen in virtually every tissue.
- Cause: Absence of the alpha-1,4-Glucosidase enzyme, which normally breaks down
polyglucose in lysosomes in all cells. This leads to obstruction of lysosomal function.
- Symptom: Ultimately leads to massive cardiomegaly (due to glycogen in cardiac
muscle) and death before 30.
- Type-III (Cori's Disease): Mild glycogen accumulation, due to a deficiency in the
debranching enzyme.. Essentially, inefficient utilization of glycogen with a little over
storage.
- Hepatomegaly occurs in early age but then diminishes.
- Relatively benign condition.
- Type-IV (McArdle's Disease): Absence of Muscle Glycogen Phosphorylase, so that
muscle glycogen stores are unavailable to exercising muscle.
- Lab: Increased levels of Muscle CPK (Creatine Phosphokinase) and myoglobin.
These symptoms generally indicated any muscle disorder.
- Very serious condition -- defective cardiac muscle.
Return to top
THE PENTOSE-PHOSPHATE SHUNT
OVERALL PURPOSE:
- It produces 2NADPH, which is used instead of NADH in most biosynthetic pathways. This
is the primary (perhaps only?) source of NADPH in metabolism.
- NADPH is needed in red blood cells as a cofactor for Glutathione Reductase, which
is essential for the RBC. RBC's can utilize the pentose phosphate shunt to supply this
NADPH.
- It produces Ribose-5-Phosphate, the essential precursor to nucleotide synthesis.
PATHWAY:
- Glucose ------> Glucose-6-Phosphate
- Catalyzed by Hexokinase; -1ATP
- Glucose-6-Phosphate ------> 6-Phosphogluconolactone
- Catalyzed by Glucose-6-Phosphate Dehydrogenase. This is a REDOX reaction,
coupled to NADP.
- This is an internal-ester -- a lactone ring.
- Concurrent NADP ------> NADPH Reduction.
- 6-Phosphogluconolactone ------> 6-Phosphogluconate -- a hydrolysis reaction.
- H2O is added across the lactone to open the ring.
- Catalyzed by 6-Phosphogluconolactase.
- 6-Phosphogluconate ------> Ribulose-5-Phosphate -- a redox reaction
- Catalyzed by 6-Phosphogluconate Dehydrogenase.
- It is the oxidation of an alcohol function to a keto group, and a decarboxylation,
leaving a 5-carbon-sugar skeleton.
- Concurrent reduction of NADP ------> NADPH
- Ribulose-5-Phosphate ------> Ribose-5-Phosphate
- This is done via an isomerase.
- The reactant can also go to Xylulose-5-Phosphate via an epimerase, but that's not
metabolically important.
- The Ribose-5-Phosphate can then serve as the precursor for nucleotide synthesis.
- Through carbon-recombination, Fructose-6-Phosphate is an ultimate product, which can then
be reconverted back to Glucose-6-Phosphate, which can then start the cycle over again!
- Each time the pentose-phosphate pathway recycles as above, you oxidize the
equivalent of one carbon from glucose. So, if you run the cycle 6 times, you
completely oxidize one glucose.
- This oxidation of glucose is an alternative to glycolysis and the TCA cycle, which is
not energetically important, but it does provide essential intermediates -- NADPH and
Ribose-5-Phosphate.
Return to top
GLUCONEOGENESIS
Q: WHAT CAN BE CONVERTED BACK TO GLUCOSE?
A: Lactate, pyruvate, or any three-carbon equivalent. Acetyl-CoA CANNOT BE CONVERTED
BACK TO GLUCOSE!
LIVER: The main organ that does gluconeogenesis.
MUSCLE: Muscle does gluconeogenesis too if need be, to create glucose-6-phosphate. However,
it cannot go all the way back to free glucose.
PATHWAY: Starting from Lactate, and going to Glucose
- Lactate ------> Pyruvate (oxidation)
- Catalyzed by Lactate Dehydrogenase
- Concurrent NAD+ ------> NADH (reduction)
- Pyruvate ------> Oxaloacetate
- The ultimate goal here is to get back to Phosphoenolpyruvate, so that we can
continue with the reverse of glycolysis. But since PEP is an irreversible reaction in
glycolysis, we need a workaround.
- This reaction catalyzed by Pyruvate Carboxylase.
- ATP is required.
- This reaction occurs in the mitochondria.
- Oxaloacetate ------> Phosphoenolpyruvate
- GTP is consumed. So, in two steps we have gotten to PEP, at the expense of ATP
and GTP, or energetic equivalent of 2ATP.
- Catalyzed by Phosphoenolpyruvate Carboxykinase.
- The malate shuttle allows free passage of this intermediate to the cytosol, so that
gluconeogenesis can be completed.
- Phosphoenolpyruvate ------> ------> Glucose-6-Phosphate
- The rest of the intermediates are the same as they are for glycolysis.
- Fructose-1,6-biphosphate ------> Fructose-6-Phosphate was irreversible in
glycolysis, so here it must be catalyzed by a different enzyme, although we still have
the same intermediates.
ENERGETICS OF GLUCONEOGENESIS: It costs 6 ATP to make one glucose -- 4 ATP + 2
GTP.
LIVER ONLY: Glucose-6-Phosphatase can catalyze Glucose-6-Phosphate ------> free glucose,
which can then be excreted into the blood to raise blood-sugar. This does not occur in muscles,
although muscles undergo gluconeogenesis.
REGULATION OF GLUCONEOGENESIS:
- Positive Effectors: Generally things that indicate a surplus of energy.
- ATP
- Acetyl-CoA
- Citrate
- Negative Effectors: Generally things that indicate a lack of energy, as well as glycolytic
intermediates.
- AMP
- Fructose-1,6-biphosphate or Fructose-2,6-biphosphate
- Inorganic phosphate
Return to top
INTERCONVERSION OF SUGARS / OLIGOSACCHARIDES / GLYCOPROTEINS
METABOLISM OF GALACTOSE: Galactose (commonly from lactose, or milk sugar) must be
activated to UDP-Galactose before it can be metabolized. Once activated, it can be freely
interconverted with UDP-Glucose for further metabolism. Starting with free galactose:
- Galactokinase: Galactose + ATP ------> Galactose-1-Phosphate + ADP
- Glactose-1-Phosphate Uridyltransferase: Galactose-1-Phosphate + UDP-Glucose
------> Glucose-1-Phosphate + UDP-Galaactose
- Galactose-4-Epimerase: UDP-Galactose <====> UDP-Glucose
- This is a two step reaction requiring NAD+ as a cofactor, although there is no net
change in NAD+.
- Step I: Oxidation of the OH at the position to a carbonyl.
- Step II: Reduction of the OH at the 4 position, to change its direction.
GLUCURONIC ACID: Important derivative of glucose, with the C6 carbon oxidized all the way
to a carboxylic acid.
- It is used in the conjugation of drugs in the liver, to make them soluble so they can be
excreted in the urine.
- It is a carboxylic acid and thereby negatively charged, enhancing its ability to
solubilize.
- It is an important component of hetero-polysaccharides.
- SYNTHESIS = a double-oxidation: C6-OH ------> C6-Aldehyde ------> C6-Acid
- Concurrent Reduction: 2 NAD+ ------> 2 NADH
GLUCOSAMINE: The acetylated form is an important component of glycoproteins. Unacetylated,
it constitutes the exoskeleton of insects.
- SYNTHESIS: An amino group is transferred from Glutamine. Then the product is
acetylated.
- Fructose-6-Phosphate ------> Glucosamine-6-Phosphate
- Concurrent loss of NH3: Glutamine ------> Glutamate
- Glucosamine-6-Phosphate ------> N-Acetylglucosamine
N-LINKED GLYCOSYLATION: An N-Glycosyl-Link is formed between N-Acetylglucosamine
and an Asp residue.
- CONSENSUS SEQUENCE: Asn-X-(Thr or Ser). This is the consensus sequence found
on almost all proteins where N-Linked Glycosylation might occur.
- X can be any residue except Proline.
- There is a CORE of two N-Acetylglucosamines and 2 mannoses that are hooked to the Asn
residue.
- 20-50 more sugars are attached. Most sugars are found in the chains, but typically not N-Acetylgalactosamine.
- N-Linked sugars are stable in alkaline environment.
O-LINKED GLYCOSYLATION: N-Acetylgalactosamine is the more common sugar. It is
attached via a hydroxy group to a Ser or Thr residue.
- O-Linked sugars are unstable in alkaline environment.
- The sugar chain can be hundreds of sugars long.
- O-Links often contain N-Acetyl-Neuraminic Acid and Fucose, but never Glucuronic Acid.
- Linked by 1,2 and 1,3 linkages to each other.
HYDROXYLYSINE: Found in collagens. Glucose residues, in groups of only 2 or 3, are
commonly glycosylated to Hydroxylysine residues on collagen-chains.
DOLICHOL PHOSPHATE: Aids in N-Linked glycosylation in the Golgi Membrane (or associated
with it).
- Dolichol Phosphate hooks up with charged UDP-N-Acetylglucosamine, to form a sugar with
a long fatty tail. Then another residue of the same thing comes.
- Then multiple mannoses are added, charged in the form of GDP-Mannose.
- In the end, we have a sugar core of a series of mannoses, and two N-Acetylglucosamines.
- In this form, the Dolichol Pyrophosphate complex transfers the sugar groups to a suitable
protein residue.
HETEROPOLYSACCHARIDES: Found in the extracellular matrix, synovial fluid, lubricants, etc.
- CHONDROITIN SULFATE: A polymer of glucuronic acid and N-Acetylgalactosamine
sulfate.
- Hydroxy group at C4 of the N-Acetylgalactosamine is sulfated.
- It is a highly charged polymer, found in ground substance, having a high water of
hydration.
- HEPARIN: Heteropolysaccharide -- natural anti-coagulant.
- A disaccharide of glucuronic acid and N-Acetylglucosamine-6-Phosphate.
- It is extremely negatively charged with one carboxylate and three sulfates.
- Produced in high quantities in the lungs.
Return to top