
Taxonomy and Phylogeny
Taxonomy
Dinoflagellates
belong to the Kingdom Protista: Division Pyrrophyta (Taylor 1987, Dodge 1985).
Dodge recognizes only one class, Dinophyea, while Taylor does not include any
classes in his index of taxonomy. The number of orders varies by publication.
Dodge (1985) identified 6 orders, 17 families, and 31 genera. Taylor (1987)
provides a more extensive review which includes 14 orders, 49 families, and more
than 150 genera. The number of dinoflagellate species is estimated to be greater
than 2000 (Barby
2001) With the recent advances in DNA and rRNA identification, these numbers
are likely to increase, if they have not done so already.
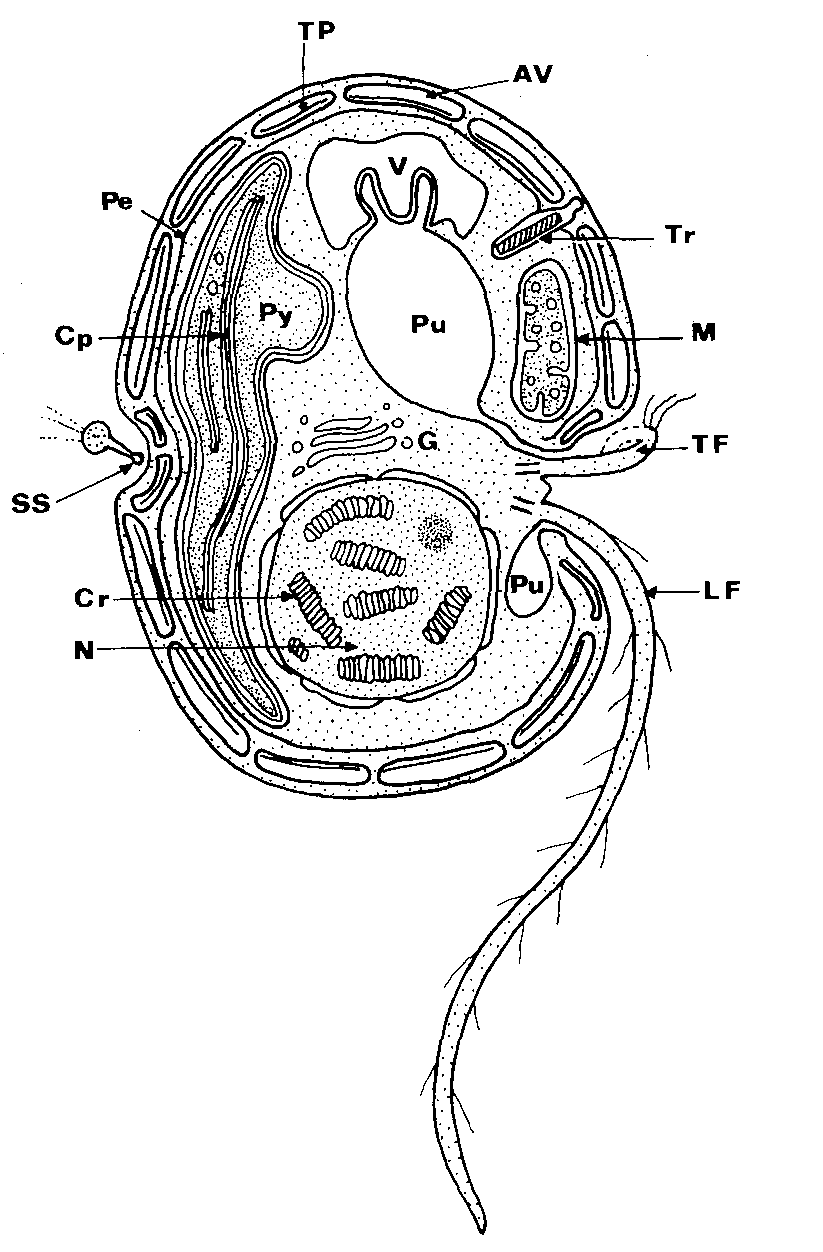
Because of their unique characteristics, dinoflagellates have been described as both plants and animals. This is because about half of the more than 2000 described species are autotrophic, and the other half is heterotrophic (Barby 2001). Classification of dinoflagellates has been very difficult because there is no one feature that is common to all dinoflagellates (Taylor 1987). Figure 1 from Taylor (1987) is a cross sectional view of a dinoflagellate; features that are important for classification are labeled. Fossil forms are most commonly identified using cyst morphology while modern, living forms are often classified based on thecal morphology. This, of course, leads to problems linking extant and extinct organisms of the same species.
Figure 1. Cross sectional view of a dinoflagellate with important features labled (Taylor 1987)
Living species are identified using several criteria (see Table 1). These include: nuclear structure, flagella, thecal plates, organelles, trichocysts, size, girdle position, life cycle, and many others.
| Character | Species | Genus | Higher |
|
|
Size |
+ |
|
|
|
|
Form |
+ |
+ |
+ |
|
|
girdle position |
+ |
+ |
|
|
|
displacement |
+ |
+ |
|
|
|
Tabulation |
+ |
+ |
+ |
|
|
girdle plates |
|
+ |
+ |
|
|
antipicals |
|
+ |
+ |
|
|
Ornamentation |
+ |
|
|
|
|
Plastid presence |
+ |
|
|
|
|
pyrenoid number |
|
1 |
|
|
|
Biochemical |
3 |
2 |
|
|
|
Life cycle |
|
|
|
|
|
phase dominance |
|
+ |
+ |
|
|
cyst morphology |
+ |
+ |
|
|
|
|
|
|
|
|
|
1 Usually not use
as a generic criterion in dinoflagellates |
||||
|
although commonly
used at this level in other groups. In |
||||
|
Pheoploykrikos
its distinction form Polykrikos is bolstered |
||||
|
by the nuclear
membrane. |
|
|
|
|
|
2 The number of
pyrenoids has been used to distinguish |
||||
|
the coccoid,
symbiotic genera Symbiodinium and |
|
|||
|
Zooxanthella,
although not all authors agree. |
|
|||
|
3 Features such as
toxins and luminescence have sometimes |
||||
|
been used as
primary characteristics, e.g. in Protogonyaulax, |
||||
|
but have been rejected as unreliable or impractical later. Table
1. Common characteristics used to classify dinoflagellates (Taylor
1987) |
||||
Phylogeny
The phylogeny of the dinoflagellates is rather vague; it is commonly determined by examining two prominent characteristics: the dinokaryon and the amphiesmal (thecal) vesicles (Saldarriga et al. 2001). The dinokaryon is a modified nucleus that is unique to dinoflagellates. It contains no histone proteins and the chromosomes remain condensed and attached to the nuclear envelope (Saldarriga et al. 2001, Sze 1998). The amphiesmal vesicles lie under the cell membrane; in some dinoflagellates, these vesicles contain the thecal plates (Speer and Waggoner 2002).
The method of using thecal plates to classify dinoflagellates is called tabulation. Tabulation has been used to determine relationships up to the family level, but higher orders of classification using this method are not possible (Taylor 1999). Two distinct types of dinoflagellates can be discerned by the size and number of thecal plates; these are the desmokonts and the dinokonts). Desmokonts usually have two large plates, while dinokonts have variable thecal plates. These two types are also defined by the location of the flagella (Sze 1998).
There are problems that arise from the use of thecal plates to determine the evolutionary history of dinoflagellates. This is because phycologists working on this matter do not agree on which form is the more primitive morph. Some argue that primitive forms are those with several small plates, and that, over time, the pates have grown in size and decreased in number (Sze 1998). Others argue just the opposite; they suggest that the primitive morphs had few plates and that the number of plates has increased over time. This model is illustrated in Figure 2. Athecate forms also pose a problem because their evolutionary history is not able to be determined by the size, shape, and number of thecal plates (Taylor 1999).
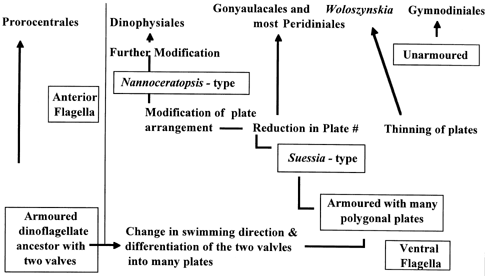
Figure 2. Evolution of Dinoflagellates based on paleontological data (Drayton 2002)
Recently, ribosomal RNA has been used to identify the relationships between dinoflagellates and other groups of organisms, and within the dinoflagellates themselves (Sze 1998, Saldarriga et al. 2001). Although relatively few dinoflagellates have been sequenced, researchers have determined that they are closely related to the ciliates, but more closely related to the apicomplexans (Saldarriga et al. 2001, Sze 1998, Sogin and Patterson 2002). Figure 4 depicts the phylogenetic relationships between dinoflagellates and other closely related organisms.
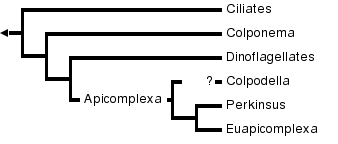
Figure 4. Phylogenetic tree relating dinoflagellates to apicomplexans and other Protista (Sogin and Patterson 2002)
Page created by: Amanda Windsor