Derek Wong
Neuromuscular Lab Report
Introduction
Muscles are different from other tissues in that it has the
ability to transform chemical energy, in the form of ATP, into
directed mechanical energy. This allows muscles to be
capable of exerting force.
There are three types of muscles, skeletal, cardiac, and
smooth. This experiment
will concentrate on the skeletal muscles, specifically the
gastrocnemius muscle, in the Bufo marinus and examine the
properties of the neuromuscular unit. The neuromuscular unit focused
on will be the sciatic nerve controlling the gastrocnemius
muscle. These properties include
threshold, maximum threshold, parts of muscle contraction, temporal
summation, minimum delay of stimulus to delay summation, effect of
varying degrees of stimulus, twitch, tetanus, and
fatigue.
The sciatic nerve is the longest and thickest nerve in the
toad. It supplies the entire lower
limb, except the anteromedial
thigh. The sciatic
nerve actually consists of two nerves, the tibial and the fibular,
which are wrapped, in a common sheath. The tibial nerve is the one
that actually innervates the
gastrocnemius. The
gastrocnemius muscle is described as a superficial muscle that forms
the proximal curve of the calf. The origin of the gastrocnemius is at
the two heads of the medial and lateral condyles of the
femur. The insertion is
by the posterior calcaneus via the calcaneal tendon (Marieb
2000).
The
functional characteristics of muscles are excitability,
contractility, extensibility, and
elasticity. Excitability
is the ability to receive and respond to a stimulus. A stimulus is an environmental
change and may arise from either inside or outside the
body. In this particular
experiment, varying degrees of electrical stimuli will be applied to
the sciatic nerve, which will then stimulate the gastrocnemius muscle
in the toad.
Contractility is the ability to shorten when adequately
stimulated. Extensibility is the ability
to be stretched. When
muscles are contracting, muscle fibers shorten, but when relaxed the
muscle fibers can relax beyond resting
length. Elasticity is the
ability of a muscle fiber to recoil and resume resting length after
being stretched. (Campbell 1998)
Muscle fibers in vertebrate muscles are organized into
motorunits. The processes of a single
nerve cell, called a motor neuron, by branching, innervating several
muscle fibers. The cell body of a motor neron
is locatd in a ventral horn of the spinal cord, and its axon ends on
themuscle fiber in a specialized region called the motor end
plate. Surrounding the
muscle cell is a sheath composed of a sheath of membrane material and
reticular fibers.
Immediately under this is teh muscle cell membrane, which is
called the sarcolemma or the plasmalemma (Carlson,
1975).
In
order for a muscle to be able to exert any force, it must first be
stimulated. Skeletal muscles are
stimulated by motor neurons of the somatic (voluntary) nervous
system. When a motor neuron transmits
an electrical impulse, all of the muscle fibers that it innervates
respond by contracting. Once a nerve impulse reaches the end of an
axon, the voltage-regulated calcium channels open and allow calcium
to flow into the extracellular fluid. The presence of calcium causes
the synaptic vesicles to fuse with the axonal membrane and release
the neural transmitter, acetylcholine (ACh). The ACh then binds to ACh
receptors on the motor end plate, which then opens the sodium
channels and initializes the depolarization of the sarcolemma (Eckert
1997).
At rest, the electrical condition of the polarized sarcolemma
is such that the outside is positive, while the inside is
negative. The predominant extracellular
ion is sodium, and the predominant intracellular ion is
potassium. At this stage the sarcolemma
is relatively impermeable to both
ions. When stimulation
occurs, by the release of ACh, the sodium channels on the sarcolemma
open and allow sodium to enter.
As sodium ions enter the cell, the resting potential is
increased, thus causing
depolarization. If the
stimulus is strong enough, an action potential is initiated. This positive charge on one
section of the sarcoplasmic retiuculum changes the permeability of
the adjacent section, causing an opening of voltage-gated sodium
channels to open there. That section's membrane
potential will then decrease and depolarization will occur there as
well. This action potential will
travel rapidly over the entire
sarcolemma. Immediately
after the depolarization, the sarcolemma's permeability changes such
that sodium channels close, but potassium channels open, allowing for
potassium to diffuse from the
cell. With potassium
leaving the cell, the electrical conditions are restored. Repolarization occurs in the
same direction as the
depolarization.
Repolarization must occur in order for the muscle fiber to be
stimulated again. If repolatization does not
occur, depolarization can not take
place. The ionic
concentrations of the resting state are later restored by the
sodium-potassium pump (Davis 2001).
Once the action potential is generated by the motor neuron, it
is propagated along the sarcolemma and down T
tubules. The action
potential then triggers the release of calcium from the terminal
cisternae of the sarcoplasmic
reticulum. The calcium
then binds to troponin, which causes troponin to undergo
conformational changes.
This conformational change removes the blocking action of the
tropomyosin, resulting in the actin active site to be
exposed. The myosin cross
bridges then alternately attach to actin and detach, pulling the
actin filaments towards the center of the sarcomere. The release of energy by ATP
hydrolysis powers the cycling process. When the action potential
ends, the calcium is removed by active transport and pumped back into
the sarcoplasmic reticulum and the muscle repeats the cycle (Hodkin
2000).. Contraction continues until the calcium signal
ends. When this happens,
the tropomyosin returns to its original location to block the actin
active site, causing the contraction to end and muscle fibers to
relax (Huddart 1997).
It
is important to note that the muscle may not shorten when
stimulated. In isotonic (concentric)
contractions, the muscle changes in length when
stimulated. The isotonic
muscle shortens and does work. Isometric (eccentric) contractions are
opposite of isotonic muscles in that when stimulated the muscle does
not shorten to do work. Although skeletal muscles are described as
"voluntary muscles," even relaxed muscles are almost always in a
slightly contracted state.
This phenomenon is called muscle tone. Muscle tone does not
produce active movements, but keeps the muscles firm and ready to
respond to stimulation (Marieb
2000).
The response of a muscle may be separated into different
stages. The latent period is the first
few milliseconds following stimulation when excitation-contraction is
occurring and there is an increase in muscle
tension. This is the
period from action potential initiation to the beginning of
mechanical activity. The next period is contraction. Muscle contraction is defined
as the generation of force (tension) by the myosin cross bridges. The
last phase, period of relaxation, is initiated by the reentry of
calcium into the sacroplasmic
reticulum. Since
the contractile force is no longer being generated, muscle tension
decreases to zero, and the tracing returns to
baseline. If the muscle
had shortened during contraction, it now returns to its initial
length (Campbell 1997).
A muscle twitch is the response of a muscle to a single brief
threshold stimulus. When a single stimulus is delivered, the muscle
contracts and relaxes. Graded responses are the
various degrees of muscle
contractions. Muscle
contractions can be graded by either changing the frequency (speed)
of the stimulation or by changing the strength of the
stimulus. When two
identical stimuli are delivered to the muscle in rapid succession,
the second twitch will be stronger than the first. This phenomenon is called wave
or temporal summation and on the physiograph the second spike
(twitch) will be shown to be higher than the
first. This type of
summation occurs because the muscle has not had time to completely
relax and since the muscle is already partially contracted, the
tension produced during the second contractions causes more
shortening than the first so the contractions are
summed. Incomplete
tetanus is when the stimulus is held at a constant strength or
voltage, and the time for muscle relaxation becomes shorter and
shorter. This will cause the degree of
summation to be greater and greater and lead to a quivering
contraction. Complete tetanus is a smooth,
continuous contraction without any showing of relaxation (Bayliss
2000).
In
the environment, there are constant
stimuli. However, not all
stimuli will cause muscle contractions. A certain level of stimulus
must be reached in order for a muscle contraction to be
achieved. The stimulus at
which the first observable muscle contraction occurs is called the
threshold stimulus. Once
past the threshold stimulus, the muscle contracts more and more until
maximal stimulus is reached.
Maximal stimulus is the strongest stimulus that produces
increased contractile force. If stimulus intensity is
increased beyond this point, the muscle contraction does not become
stronger (Carlson 1974).
Regardless of the amount of stimuli, muscle activity cannot continue indefinitely. Fatigue is a situation in which the muscle is unable to contract and tension drops to zero. In this experiment, fatigue can also be defined as any decline in force output during prolonged stimulation. The reason this occurs is because during fatigue there is a substantial effux of potassium ions that allows for an increase in extra-cellular potassium concentration, causing the depolarization of the membrane. This then depolarizes the membrane inactivating the sodium channels and reducing membrane excitability. In short, the fatigue caused by the repeated stimuli will result in the decrease of the muscular response (Campbell 1998). This response is a phenomenon called the Wedensky inhibition.
Methods and Materials
Preparation of the Gastrocnemius
A
double pithed Bufo Marinus was obtained, and its leg was
pinned to the dissection tray to prevent movement. The gastrocnemius muscle was
then isolated and fastened by a strong thread around the achilles
tendon. The tendon was then cut free
of its attachment to the heel and tied to the other end of the thread
to the transducer (Grass FT
103). The transducer was
placed above the gastrocnemius muscle and held in place by a clamp
connected to a metal stand.
The transducer was then connected to the preamplifier (Grass
PI-5), which was then connected to the physiograph (Grass
700).
The skin from along the length of the leg was removed to
reveal the large muscle groups of the thigh and
calf. The muscles on the
medial sides were teased apart and the sciatic nerve, located along
the anterio-medial side of the femur, was
found. The sciatic nerve
was then lifted up gently by the glass probe and attached to the
sleeve electrode. The
sleeve electrode was then connected to the stimulator (Grass SD9).
Both the gastrocnemius muscle and the sciatic nerve was constantly
kept hydrated by the Normal Ringer's solution throughout the duration
of the experiment.
Twitch Threshold
The paper speed was first set to 1 mm/s and the stimulator
voltage intensity was set to zero with duration of 2 ms. The nerve
was then stimulated by a single impulse between five second
intervals. The impulse
continued as the voltage intensity increased from zero until no there
was no increase in the twitch height observed. After this procedure, the
force of contraction was calculated for each trial using the
following equation:
Force of contraction (in
grams) = (Peak in amplitude)*(Calibration constant in
gram/cm)
Voltage Duration
The paper speed was changed to 2.5 mm/sec and the stimulator's
voltage was altered so that the twitch height response was equal to 1
centimeter. The stimulus voltage and
duration was then changed so that for each different combination of
voltage and duration, the twitch response is 1
cm. This was done for
five different trials, such that no two trials had the same stimulus
intensity.
Phase Contraction
In this part of the experiment, the paper speed was set to the
fastest setting (50 mm/s).
The purpose was to measure the latency period, the duration of
contraction, and the duration of relaxation of a
twitch. The stimuli were
applied by pressing the stimulator button and the event marker
simultaneously.
Summation of Subliminal Stimuli
The
threshold voltage was reestablished by using manual single shocks
with duration of 2 milliseconds.
The stimulus intensity was then set just below the threshold
voltage. Here, the stimulus intensity was set to 0.32 V for duration
of 2 milliseconds. The single stimuli
switch on the stimulator was pressed 5 times in rapid
succession.
Effect of frequency of stimuli (induction of tetanus)
The paper speed was adjusted to 1 mm/s and adequate stimulus
strength was chosen from part one. Here, the stimulus was set to
0.33V. The nerve was then stimulated
for about five seconds on "repeat mode" at various stimulator
frequencies starting at 2 per second, increasing frequency until
tetanus (continual contraction) is observed.
Minimum Stimulus delay to induce summation and/or
refraction
The stimulator was set to "twin pulses," and in single mode
the voltage intensity was set to produce a response of 1 centimeter
over a duration period of 2
milliseconds. Here, the
intensity was set to 0.95 volts. The delay knob on the stimulator was
used in order to vary the time between the twin impulses. The delay
times used were 200 milliseconds, 100 milliseconds, 20 milliseconds,
10 milliseconds, 1 milliseconds and 0.1 milliseconds.
Fatigue
The paper speed was set to 1 mm/s and the stimulator frequency
was set to 25 per second.
An adequate stimulus intensity, 8 volts, was
chosen. The nerve was
stimulated until fatigue was observed and a decrease in the amplitude
of the stimulus. The
stimulus voltage intensity was continuously raised until the response
was completely baseline. The stimulator was then turned
off and the nerve sleeve electrode was
disconnected. The
stimulator was then turned back on, and the muscle was stimulated
directly by placing the two-pin electrode directly on the belly of
the muscle. The nerve
sleeve electrodes were reconnected and the nerve was stimulated as
before. After the stimulation, a five
minutes waiting period took place in order to allow for
recovery. Once the recovery period had
passed, the nerve was stimulated again.
Calibration
The final part of this experiment was to calibrate the
physiograph. Removed the string from the
muscle and tie a weight to the string. The paper speed was set to 5
mm/s, the weight was allowed to drop, and readings on the physiograph
were observed. The calibration constant was
then calculated by dividing the weight (grams) by the defection on
physiograph (cm).
Results
1. Twitch
Threshold
Stimulus
intensity (mV) required to produce a threshold of 420 mV and
maximal 600 mV response when the duration is 2
ms.
Table 1: The Peak Amplitude and Force of Contraction for different Stimulus Intensities
|
Stimulus
Intensity (V) |
Peak
Amplitude (Height in cm) |
Force
of Contraction (g) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
According
to Table 1, as the stimulus intensity increased, the peak amplitude
increased. As the peak amplitude increased, so did the force of
contraction. The stimulus intensity
increased from 0.46 volts to 0.68 volts, the peak amplitude increased
from 1.00 cm to 2.80 cm and the force of contraction increased from
7.143 grams to 20.00 grams. When the stimulus was
increased from 0.64 volts to 0.66 volts, there was no difference in
the peak amplitude and the force of contraction.
<![if !vml]>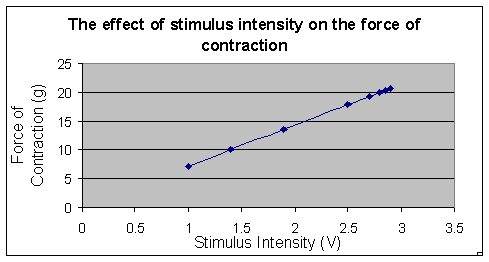 <![endif]>
<![endif]>
Figure 1:
The effect of stimulus intensity on the force of contraction of a
gastrocnmeius muscle of a Bufo
marinus. The above graph
shows pictorially the force of contraction on the y-axis and the
stimulus intensity on the
x-axis. As shown on the
graph, as the stimulus intensity increase so does the force of
contraction.
2.
Voltage/Duration
Table 2:
The Effect of Duration on Threshold Stimulus
Intensity.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
According
to Table 2, as the stimulus intensity decreased from 0.48 volts to
0.4 volts, the duration increased from 2 ms to 8
ms.
<![if !vml]>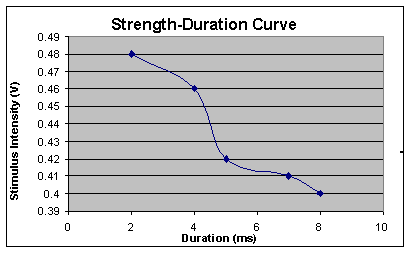 <![endif]>
<![endif]>
Figure 2:
Strength-Duration Curve. The above graph shows pictorially the
stimulus on the y-axis and the duration on the
x-axis. As shown on the
graph, as the stimulus intensity increases the length of duration
decreases.
3. Phase
of Contraction
<![if !supportLists]>A.
<![endif]>Latency Period:
20 ms
<![if !supportLists]>B.
<![endif]>Contraction
Period: 30 ms
<![if !supportLists]>C.
<![endif]>Relaxation Period:
150 ms
Duration: 2 ms
Intensity: 0.32 volts
Paper speed: 50 m/s
There are
three different durations for each of the different periods during
the phase of contraction.
The latency period was the shortest being only 20 ms, followed
by the contraction period which was 30 ms, and the relaxation period
was the longest being 150 ms.
<![if !supportLists]>4.
<![endif]>Figure 4:
Summation of Subliminal Stimuli - do I have to include anything
more?
This
graph shows five
rapid
succession of a
stimuli
below the
threshold previously
determined(at
0.42 volts).
As more
stimuli was
applied,
peak amplitude
increased.
<![if !supportLists]>5.
<![endif]>Effect of
frequency of stimuli (induction of
tetanus).
Figure 5.
This physiograph shows the effect of a frequency of 14 pulses on a
motor unit. Paper speed was 25 mm/s, the voltage intensity was 3
volts and the duration was 2
milliseconds. This graph
shows that as the stimulation was constantly applied, the muscle
twitched with a greater force.
Eventually it remained at a steady peak
amplitude.
<![if !supportLists]>6.
<![endif]>Minimum stimulus
delay to induce summation and/or refraction
Table 3: Determination of minimum stimulus to induce wave summation
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Table 3
shows that when the delay between twin pulses was 20 ms or lower,
there were no longer separate peaks, summation occurred and there was
no stimuli observed during the refractory
period. When the delay
was decreased to 1 second and 0.1 seconds, the peak observed was
substantially smaller than when the delay was 2 ms.
Figure 6:
Minimum stimulus delay to induce wave summation.
Once the
delay between twin pulses was less than 100 ms, there was no stimuli
noticed during the refractory
period.
<![if !supportLists]>7.
<![endif]>Figure 7:
Fatigue. This graph shows fatigue of
the motor unit in the gastrocnemius muscle of the Bufo marinus after
constant stimulation of 8 volts with a frequency of 25 pulses and
duration of 2 milliseconds. Paper speed was set to 2.5
mm/s.
The peak amplitude increased sharply in the beginning of stimulation and then remained at a constant level for the duration of the stimulation.
<![if !supportLists]>8.
<![endif]>Calibration
Table 4:
Calibration
|
Weight
(g) |
Deflection
on Physiograph (cm) |
|
5
grams |
0.7
cm |
|
10
grams |
1.25
cm |
This
table shows the values obtained using different weights to determine
the ratio that was used to solve for the force of
contraction.
Figure 8:
Post-fatigue stimulation. This graphs shows that direct stimulation
was applied to the gastrocnemius muscle of the Bufo marinus after the
muscle was fatigued in the previous parts of the
experiment. The muscle
showed that it could still exert force. The voltage was 3 volts, and
the paper speed was 2.5 mm/s
Discussion
In
this experiment, the gastrocnemius muscle was stimulated in a
multitude of ways and observations were
taken. In the first
experiment, the twitch threshold and maximal contraction was
determined. The voltage and duration at which a twitch was first
observed was the twitch
threshold. Threshold was
at 420 mV. The point at which the twitch
height no longer increased was the maximal
contraction. The maximal
contraction was 600 mV for a duration of 2 ms. This maximal
contraction rate is determined by the rate at which cross-bridges
detach from the actin thin filaments (Eckert
1997). At maximal
contraction, all of the possible myosin cross-bridges in the muscle
has detached. This is the
point at which all of the muscle's motor unit has been
recruited.
The recruitment of motor units is a phenomenon when activating
more of the motor neurons that control the muscle can progressively
increase the tension in a
muscle. Depending on the
task at hand or stimulation being administered, the brain recruits
the appropriate number and type and size of the motor
neurons. For delicate
jobs, the brain will most likely use much fewer motor neurons,
compared to a task that involves heavy lifting (Campbell
1998).
The second half of the first experiment had the stimulus
increased over time. It was observed that as the stimulus increased
the force of contraction increased as
well. This shows that as
the stimulus intensity increases the force of contraction increases.
This direct correlation is due to the fact that nerves undergo an
all-or-none event. This phenomenon simply states
that once an action potential is either reached and then perpetuated
down the nerve, or there is no action potential at
all. The sciatic nerve,
as well as many other multi-fiber preparations, is actually a
substantially large bundle of
nerves. The more nerves
that are stimulated, the greater the force of
contraction. The nerves
in these large bundle of nerves have threshold voltages that cover a
wide range. As stimulus intensity
increases, more individual nerves are
stimulated. As more
nerves are stimulated the muscle contracts with an even greater
force. In graph 1, it is shown that as stimulus intensity increased,
the force of contraction increased as well, supporting the idea of
more stimulus will result in more contractile muscle
force. As stimulus
increased, however, the force started to increase only by a small
amount. For example, when the stimulus
intensity was at 0.58 volts, the force of contraction was 19.29
grams. At a stimulus intensity of 0.6
volts, the force of contraction was 20.7
grams. At 0.6
volts and at 0.62 volts there was no change in the force of
contraction. This was
because all the nerves were already stimulated and the muscle was at
the maximal contraction, here it was 0.6
volts. When additional
stimulus, the force of contraction did not rise, in fact the force
decreased. This was most likely due to muscle fatigue, therefore the
muscle did not respond with as much force as in the previous parts.
This is why the graph in Figure 1 was not a hyperbolic curve but a
straight line instead. It should have been a
hyperbolic curve because as the muscle reaches the maximum threshold,
the rate at which the muscle exerts force starts to slow down
(Gozariu 1997).
The second part of the experiment dealt with duration vs.
stimulus intensity. As shown in table 2, as
stimulus intensity decreased the duration
increased. In graph
2, it shows that duration and stimulus intensity has an inverse
non-linear relationship.
This means that for long shocks, the applied current reaches a
miniscule minimum. When the duration is reduced,
a much stronger shock is necessary to reach the same threshold
(Marieb 2000).
The
longer the duration, the more opportunities of stimulation the muscle
has to contract. This is
the same as summation, where even if the stimulation is of a
sub-threshold level, when given enough time, the muscle may reach
threshold through summation. Similarly, if the voltage of
stimulation was of significant intensity, then the muscle will not
need as much time to become
stimulated. The
sarcolemma will be easily depolarized and will not need the
additional time. Since there is an inverse
non-linear relationship between duration and intensity, the graph in
Figure 2 should have been a parabolic curve instead of what it
resulted in. Most likely the muscle was
fatigued causing the results to be
distorted.
In the third part of the experiment, the phases of
contractions were examined.
The first was the latent
period. This was the
shortest and lasted only 20ms.
This phase is the time after the initial stimulus and the
beginning of the contraction. This is when the action
potential is moving down the axon and arriving at the neuromuscular
junction. During this period, muscle
tension is beginning to increase, but no response is seen on the
physiograph (Marieb 2000).
At the neuromuscular junction, the action potential causes
acetylcholine to be released from the pre-synaptic pathways and bind
to the post-synaptic receptors, initiating the second phase,
contraction. This is the
second longest period being 30
milliseconds. The period
may last from 10 - 100 ms. This is when the cross bridges are active,
from the onset to the peak of tension development, and the
physiograph tracing rises to a
peak. If the tension
(pull) becomes great enough to overcome the resistance, the muscle
shortens. The muscle fibers are shorten due to the sliding of thick
and thin filaments of the muscle, the contraction period ends and the
relaxation period begins. The final phase, lasting 10-100 ms, is
initiated by the reentry of Ca2+ into the sacroplasmic
reticulum. Since the
contractile force is no longer being generated, muscle tension
decreases to zero (Campbell
1998).
Summation
was the fourth part of this
experiment. Temporal or
wave summation is the phenomenon in which additional stimuli cause
the peak amplitude to increase.
This means that there is a summation in the force of
contraction being exerted. When two or more stimuli are applied
almost immediately after each other, the peak amplitude will increase
causing the muscle to exert more
force. During a single
stimulation, only a small amount of the motor neurons located in the
muscle are firing. When additional stimuli is
applied, more motor neurons are
fired. With more and more
stimuli, more and more motor neurons are fired causing summation to
occur (Gozariu 1997).
Eventually, however, summation will cease to increase when all
of the motor neurons are firing.
Here,
the stimulus intensity chosen, 0.32 volts, was lower than the
threshold. The stimulator was pressed
five times and temporal summation was
observed. This was
observed because the muscle was already partially contracted. By adding additional stimulus
to an already contracted muscle, the muscle will contract with an
even greater force until the muscle becomes fully
contracted. In Figure 4,
stimulus number one is a slight contraction, and as additional
stimulus is applied, the peak amplitude substantially increases. Stimulus five is quite higher
when compared to number one showing that a muscle's strength can be
increased though the addition of stimuli to an already partially
contracted muscle.
The fifth part of the experiment was the effect of the
frequency of stimuli. As the number of stimuli increased, the peak
amplitude increased, similar to what happened in the summation part
of this experiment.
Although additional stimuli does increase the force of
contraction, the muscle does have a finite amount of contractile
power. As the number of stimuli
increases, the amount in which the force of contraction increases
start to become smaller.
This was because the carrying capacity of the muscle was
almost reached (Huddart 1975).
Figure 6 and Figure 7 are different in that there were more
stimuli applied within the same time
period. This caused the
gradation of peak amplitude to be increased
drastically. The end peak
amplitude was substantially greater than the end peak amplitude of
figure 6. This shows
again that more stimulation will cause an increase in the force of
contraction.
Fatigue was shown to be achieved in figure 8. The smooth line shown in figure 8 was caused by a series of muscle action potentials at a high frequency. When the muscle is contracted at a fast enough frequency, tetanus is achieved. Tetanus is defined has the maximal, sustained contraction of a skeletal muscle. This is caused by a high frequency of action potentials caused by continual stimulation (Campbell 1998). As the muscle reaches tetanus, the increase between each progressive peak amplitude decreases, and then eventually the peak starts to round off and merges into a smooth line. When the curve smoothed out to be a line, all of the muscle fibers in that motor unit were firing (Huddart 1975).
The sixth part of this experiment was twin
pulses. Here, the
stimulator was set to fire two consecutive pulses. The time between the two
pulses was varied and observed.
In the first pulse, the delay was 200 milliseconds. In this
case, there were two distinct peaks. There was no summation and no
stimuli occurring during the refractory
period. This means that
the muscle did have time to relax in time for the second stimulus to
take affect and cause a separate contraction (Marieb 2000). The second trial shortened the
delay time in half. There
were still two separate peaks, and no summation occurred, however a
stimulus did occur during the refractory period. This means that
because the delay time was shortened, the muscle did not have enough
time to fully relax.
Since the muscle was partially contracted when the second
stimulus was applied, the second contraction was shown as a peak in
the refractory period of the first stimulus. However, in Figure 4
there are two peaks seen for 200ms and 100ms meaning that the second
pulse happened after the muscle recovered from stimulation. Then the
delay was set to 20 and 10ms. Here, the two peaks merged
into one peak with greater
amplitudes. This is
because summation occurred, similar to part 4, and the peaks
increased (Marieb 2000).
However, during the time delays between the twin pulses of 1
and 0.1 milliseconds, the peak amplitude became
shorter. This
was because the absolute refractory period was being met. This was because the
sacrolemma of the muscle was not fully
repolarized. If complete
repolarization is incomplete, no summation can take place (Marieb
2000). The period in which
repolarization is occurring and no summation will take place is
called the absolute refractory period.
Fatigue was the seventh part of the
experiment. Fatigue in
this case is the state in which the muscle force declines to
zero. In this experiment,
fatigue is seen as a physiological characteristic and not a
psychological. There are two types of
fatigue. The first, psychological, is
when the mind thinks that it is “worn out,” but the body is still
physically capable of exerting
force. The second type,
physiological, is when the body is physically no longer capable of
producing muscle contractions (Marieb
2000).
Figure 10 shows fatigue in the motor unit after
stimulation. The flat line in the beginning
indicates that tetanus has set
in. At tetanus when all
of the motor neurons are firing.
After a short amount of time, the muscle
fatigue. This is shown as
the line drops drastically into a steep, almost vertical line. Here,
the muscle goes into a state of fatigue and the force of contraction
rapidly decreases to
zero.
After fatigue has been observed in Figure 10, a direct
stimulation to the muscle was applied through the use of pin
electrodes. This was
meant to determine if the fatigue seen in the previous part was due
to the neuromuscular junction or the muscle itself. According to the physiograph
result shown in Figure 11, upon direct stimulation the muscle can
still contract. This shows that the muscle
itself was not fatigue, only the neuromuscular
junction. Therefore the
ability of the nerve to cause muscle contractions was gradually
diminished to almost zero because the neuromuscular junction did not
have enough time to recovery from the stimuli applied (Ganong
1999). This resulted in fatigue to
set in.
Conclusion
In this experiment, the gastrocnmeius muscle was exposed to an array of stimuli to observer and learn how the neuromuscular junction works, as well as its limitations.
Threshold was found to be the minimum stimuli needed to cause
a muscle contraction. The muscle contraction is
based upon the all-or-none principal where once a stimuli reached
that threshold stimuli intensity, the action potential will start and
cause the muscle to contract. As the intensity increased,
the peak amplitude increased indicated that the force of contraction
in the muscle increased as well.
However, there was a limit to the muscle’s
strength. The maximum
stimuli value was found and this was the intensity where if anymore
voltage was added, the force of contraction would not be
increased.
This experiment showed that there are three phases
contraction. The first is the latency
period, which was the shortest.
The second is the contraction period. This was when the cross
bridges are active and was manifested by the physiograph tracing
rising to a peak. The third is the relaxation
period, and was the longest. It was seen that in the relaxation
period that the force eventually reduces to zero. This was because
the contractile force is no longer being generated, muscle tension
decreases to zero.
It
was observed that upon rapid stimulation, temporal summation
occurs. This was due to the fact that
since the muscle was already partially
contracted. Since it was
partially contracted, the additional stimuli caused the muscle to
exert a greater force.
When the stimuli was applied fast enough, then tetanus set
in. This was indicated by a
flat line.
This
experiment showed that there was a minimum stimulus delay was need in
order to induce summation.
If the second stimulation was applied before the duration of
the delay, there would be no
summation. This was
because the repolarization of the sarcolemma was not yet
complete. Repolarization is the process
in which the potassium ions are expelled from the sarcolemma and its
permeability changes such that sodium channels close, but potassium
channels open and allows for potassium to diffuse out of the
cell. When potassium
leaves the cell, the electrical conditions are restored and
depolarization may take place once
again. This process does
take time, and if not given enough time, the process would be
incomplete.
Lastly,
fatigue was observed. It
was shown that fatigue was due to the neuromuscular junction and not
the muscle itself.
Works Cited
Bayliss, Douglas A.; Dong, Xiao-Wei; Feldman, Jack L.; Funk, Gregory D.; Rekling Jen C. April 2000. Physiological Reviews. Synaptic Control of Motoneuronal Excitability. Vol. 80, No. 2, pp;. 767-852
Binder-Macleod, Stuart A. and Lee,
Samuel C. K. 2000. Effects
of activation frequency on dynamic performance of human fresh and
fatigued
muscles. Journal of
Applied Physiology. 88:
2166-2175
Campbell, Neil A. 1998. Biology, Fourth Edition. The Benjamin/Cummings Publishing Company, Inc. New York. 986-992
Carlson, Francis D. 1974 Muscle Physiology. Prentice-Hall, Inc. Englewood Cliffs, NJ. 1-111
Eckert, R. 1997. Animal Physiology, Fourth Edition. W.H. Freeman and Company. New York. 143-150, 189-198, 205-212, 370-374, 394-397
Davis, Michael. Mar 01, 2001. Basic Principles of Synaptic Physiology Illustrated by Computer Model. American Physiology vol. 25. 1-12. (In "INNOVATIONS AND IDEAS")
Ganong, W.F. 1999. Review of Medical Physiology, Nineteenth Edition. Appleton & Lange. Stamford, Conn. 49-56, 63-67.
Gozariu,
Manuela; Bragard, Dominique ;
Wille, Jean-Claude ;and
Le Bars, Daniel. December 1997.
The Journal of Neurophysiology .
Temporal
Summation of C-Fiber
Afferent Inputs: Competition Between Facilitatory and Inhibitory
Effects on C-Fiber Reflex in the
Rat. Journal of
Neurophysiology. Vol. 78 No. 6 pp.
3165-3179
<![if
!supportLineBreakNewLine]>
<![endif]>
Hogdkin AL and Huxley AF. March 2000. A quantitative description of membrane current and its application to conduction and excitation in nerve. Journal of American Physiology 117: 500–544, 1952.
Huddart, Henry 1975. The Comparitive Structure and Function of Muscle, First Edition. Pergamon Press, Oxford. 155-164, 296-343
Marieb, Elaine N. Human Anatomy and Physiology. Benjamin Cummings Holoyoke Community College. 2000. 277-483.