The avian guilds of North America, along with those of Eurasia, were once the exclusive domain of Enantiornithes, the opposite birds, with their clawed wings and upside-down ankles. However, the subsequent millienia have seen true birds, Euornithes, rise into supremecy. They way has not been easy.SAMPLE TAXA:The order Coraciiformes began, as a group of generalist ground birds very much like the modern Brachypteraciidae (the ground-roller assemblage), and quickly diversified into a number of forms. Coraciiforms evolved, as most of the earth-familiar birds did, in the southern hemisphere, but quickly spread north. By the middle Eocene, the coraciiforms were arguably the most diverse of the bird orders, and certainly enjoyed a wide range, with representatives in South America, North America, and Eurasia.
The coraciiforms did not make the march toward world domination unopposed, however. Nearly all of the avian groups from home-Earth (of which Coraciiformes is one) are descended from a Gondwanan assemblage that escaped the destruction wreaked upon the Northern Hemisphere by the Chicxulub boloid. On the Specworld, where the extinction never took place, the northern bird lineages were there to meet the coraciiforms.
The Eocene strata of the Messel Shale, in Spec's Germany, clearly demonstrate the battle being waged between the birds of the north and the south. The most prevalent bird order preserved in Messel is, by far, Twitiaviformes. These bizarre birds, possibly related to the paleognaths, were probably indigenous to Europe, at the time, although now their greatest diversity is in the Southern Hemisphere. Following closely behind the twitiavians in species-count are the coraciiforms, while several extinct orders of enantiornithian birds were already loosing ground to the southern invaders.
The competition imposed by the twitiavians greatly changed the direction of coraciiform evolution during the middle Tertiary. After their upswing in the late Eocene and early Oligocene, Coraciiformes experienced a pronounced depression and went completely extinct in the Old World, but for a few strongholds in Africa and Madagascar.
For much of the remainder of the Tertiary, coraciiforms barely hung on in the Old World with many clades (hornbills, hoopoes, scimitar-bills, and cuckoo rollers, to name a few) going completely extinct or simply failing to evolve. In the New World, however, the story was different.
The twitiavians have a long history in South America, but for some reason, their penetration of North America has never been very extensive. It was in North America, then, that the coraciiforms continued to rule with little opposition. While the Old-world species died out, the American coraciiforms experienced a renaissance of diversity, radiating into many bizarre niches, most notably the piscivorous alcedinids (kingfishers) and the bizarre, wood-boring dirostrids (pickpeckers). During the Pliocene and Pleistocene, these clades spread from North America to colonize every continent but Antarctica (there are even alcedinids in twitiaviform-dominated Australia).
Meanwhile, in Africa, the Pliocene saw a new adaptive radiation of Coracii, the primitive branch of Coraciiformes supposedly relegated to obscurity by Twitiaviformes. Sub-clade Coracii, the most basal branch of the coraciiform tree, suddenly exploded across Africa and Eurasia as Parabrachypteraciidae, or nearcrows. Today, the generalist/scavenger nearcrows are ubiquitous in Africa, Eurasia, and the Americas, filling the niches that, on Home-Earth, would be occupied by the passerine corvids, the crows.
While the Americas remain center of coraciiform diversity, many of the clade's 500 known species may be found on every continent except for Antarctica.
Dirostridae
Dirostrids, the pickpeckers, are the strangest of the coraciiformes, and arguably the most sucssesful. These creatures are reletively new additions to their family tree, arising some time during the Pliocene and quickly spreading from North America to Eurasia, Africa, and (somewhat later) South America.
The pickpeckers possess a beak structure unique in Spec's birds, although they bear a superficial resemblance to the passerine akiapolaau (Hemignathus wilsoni) of Home-Earth's Hawaii. A pickpecker's upper mandible is curved and hook-like, while the lower bill forms a blunt, robust pick, similar to the beak of an RL woodpecker. Pickpeckers feed by bracing themselves against a vertical tree-trunk will powerful talons (see below) and a stiff, bristly tail. They then slowly walk about the bark, feeling with their feet for the tell-tale vibrations of a wood-boring insect larva. Upon locating food, the pickpecker opens its mouth, specialized mandibular joints allowing the bird to yawn impossibly wide, and then drives its lower bill into the wood, shattering bark to expose the hiding food. Pickpecker skulls are composed of rather spongy bone, and their necks articulate in such a way that their hammering motion is only possible on a single plane, thus avoiding the shearing forces that would pulverize the birds' brains. A pickpecker may hammer at wood at speeds of 15 to 20 kph, stripping the protective bark from above a burrowing grub that the bird may then extract with the aid of its curved, pick-like upper bill.
(Picture by Daniel Bensen)
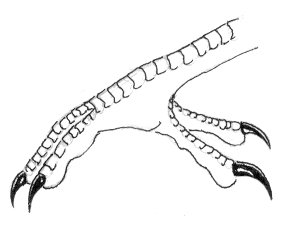
Fig 2: Zygosyndactyl footAnother interesting feature of the dirostrids' physiology is their pedal structure. Pickpecker toes are arranged in a unique 'zygosyndactyl' configuration, with digits one and four fully reversed and opposed to digits two and three, which are fused at the first joint. Zygosyndacylity is no doubt a product of the kingfishers' zygodactyl feet evolving in convergence with the syndactyl feet of the woodpeckers of Home Earth. The zygosyndactyl arrangement gives a pickpecker a firm hold on its vertical perch, and gives the bird the leverage necessary to hammer through wood.
Pickpeckers most likely evolved from insectivorous coraciiforms similar to todies or motmots (although dirostrids are considered basal alcedinoids rather than momotoids, they may be closer to motmots than kingfishers). Many motmotoids probe trees for wood-boring insects, picking away bark with their tiny beaks, and the dirostrid feeding apparatus probably evolved as an extension of this behavior.