PROCESOS CATABÓLICOS
1. ESQUEMA GENERAL DEL CATABOLISMO
En
los procesos catabólicos las moléculas orgánicas se van degradando, paso a
paso, hasta formar otras moléculas más simples y, finalmente, sustancias inorgánicas.
Globalmente son procesos de oxidación en los que las moléculas orgánicas
van perdiendo electrones que, tras pasar por una cadena transportadora, son
captados por una molécula aceptora de electrones final. En esos procesos
la energía liberada permite la formación de moléculas de ATP.
Según quien sea el aceptor final de
electrones se pueden diferenciar dos modalidades:
§
Fermentación. El aceptor final de electrones es un
compuesto orgánico, por lo que se trata de una oxidación incompleta y un
proceso anaerobio.
§ Respiración celular. El aceptor final de electrones es una sustancia inorgánica.
Si
es el O2, se trata de una respiración aerobia que realizan la
mayoría de los organismos; si es otro compuesto inorgánico (NO3-,
SO42-, CO2), se trata de una respiración
anaerobia, exclusiva de ciertos microorganismos.
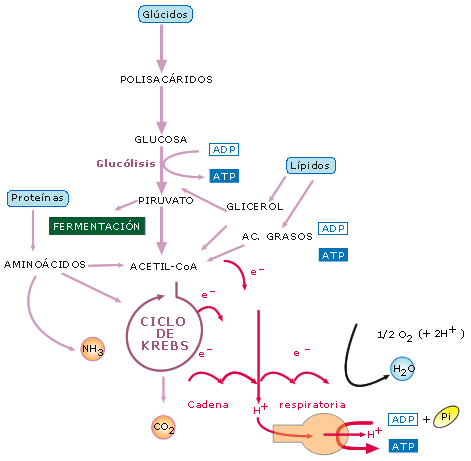
Los procesos catabólicos se pueden iniciar a partir de varios sustratos orgánicos, especialmente glúcidos, lípidos y proteínas. En cualquier caso, al final, las diferentes rutas metabólicas de la respiración celular confluyen en el ciclo de Krebs, y los electrones liberados en el proceso pasan a la cadena respiratoria para formar ATP por fosforilación oxidativa. Una variante a este esquema general es el desvío o la ruta alternativa de las fermentaciones.
Esquema del catabolismo: al tiempo que los compuestos orgánicos se descomponen en CO2 y H2O, los electrones son transferidos a lo largo de la cadena respiratoria impulsando la fosforilación oxidativa
2. EL CATABOLISMO DE LOS GLÚCIDOS
Los
glúcidos se consideran nutrientes energéticos (1 gramo de glúcidos aporta
como promedio 4 kcal). En su catabolismo, primero los polisacáridos se
descomponen hasta formar finalmente glucosa. Concretamente, en los animales, el
glucógeno acumulado en las células hepáticas o en las fibras musculares se va
hidrolizando por un proceso de glucogenolisis.
La glucosa entra a continuación en una secuencia de
reacciones.
Glucólisis
Consiste
en diez reacciones consecutivas (nueve etapas), que tienen lugar en el citosol, en las que
se diferencian dos fases:
a) Fase preparatoria. Se consumen 2 moléculas de
ATP para transformar una de glucosa en 2 de gliceraldehído 3-fosfato.
b) Fase de beneficio. Se obtienen 2 moléculas de
NADH y 4 moléculas de ATP, formándose al final de la glucólisis 2 de piruvato.
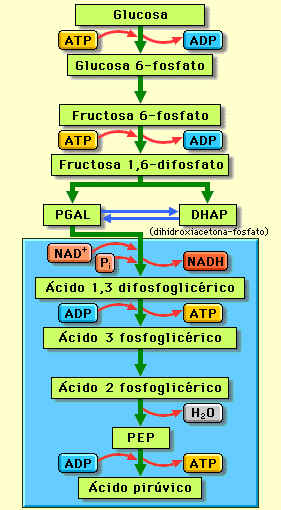
Para ver las etapas de la glucólisis en detalle pincha aquí.
En el conjunto de la glucólisis, a partir de cada molécula de glucosa, se obtienen, por tanto, como rendimiento neto 2 ATP (por fosforilación a nivel de sustrato), y 2 NADH que luego podrán pasar a la cadena respiratoria.
El siguiente esquema nos muestra el balance total de la glucólisis:
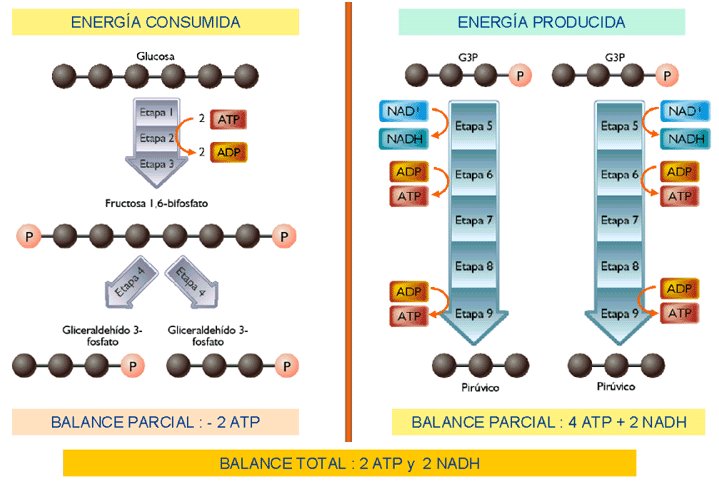
El
piruvato que se obtiene al final de la glucólisis se encuentra en un
cruce metabólico en el que puede seguir dos destinos, dependiendo de la
disponibilidad de oxígeno y el tipo de célula: la vía anaerobia de las
fermentaciones o la vía aerobia de la respiración celular.
A) Vía aerobia de la respiración celular
La
vía aerobia consiste en dos grupos de reacciones: el ciclo de Krebs, al que se
incorpora una molécula de acetil-CoA proveniente del piruvato de la glucólisis;
y la cadena de transporte de electrones, que va a servir para sintetizar ATP en
un proceso de fosforilación oxidativa.
Antes
de comenzar la vía aerobia el piruvato resultante de la glucólisis se tiene
que transformar en acetil-CoA mediante una descarboxilación oxidativa.
Descarboxilación oxidativa del piruvato
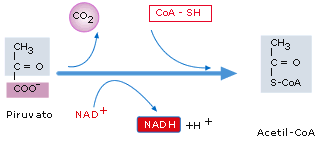 El
piruvato pasa al interior de la mitocondria, donde, mediante una reacción
irreversible, se une a un coenzima y sufre una descarboxilación (pérdida
de CO2) y una oxidación, formándose: CO2, NADH
y acetil-CoA.
El
piruvato pasa al interior de la mitocondria, donde, mediante una reacción
irreversible, se une a un coenzima y sufre una descarboxilación (pérdida
de CO2) y una oxidación, formándose: CO2, NADH
y acetil-CoA.
El
NADH liberará su poder reductor en la cadena respiratoria,
mientras que el acetil-CoA entrará en el ciclo de Krebs.
El ciclo de Krebs
Casi
todos los compuestos orgánicos (azúcares y grasas especialmente) se
descomponen hasta formar acetil-CoA: un grupo acetilo de dos carbonos,
unidos al coenzima A.
El acetil-CoA se incorpora a una secuencia cíclica de reacciones químicas que
se conocen con el nombre de ciclo de Krebs o ciclo del ácido cítrico. Este
ciclo tiene lugar en la matriz mitocondrial y consiste en una secuencia
de ocho reacciones consecutivas. En el conjunto de esas reacciones tienen lugar,
básicamente, los siguientes acontecimientos:
§
Se liberan 2 moléculas de CO2, que se puede considerar
que proceden del grupo acetilo del acetil-CoA, con lo cual se consigue una oxidación
completa de la materia orgánica. El CO2 será, por tanto, el
producto residual último de la respiración aerobia.
§
Los electrones más los protones (hidrógeno), que pierde el grupo
acetilo al oxidarse, van a parar, en cuatro fases, a transportadores de
electrones, formándose 3 moléculas de NADH + H+ y una molécula
de FADH2. El poder reductor de estas moléculas será transferido
luego a través de la cadena respiratoria mitocondrial.
§ En un paso del ciclo tiene lugar una fosforilación a nivel de sustrato que origina una molécula de GTP (equivalente desde el punto de vista energético a un ATP).
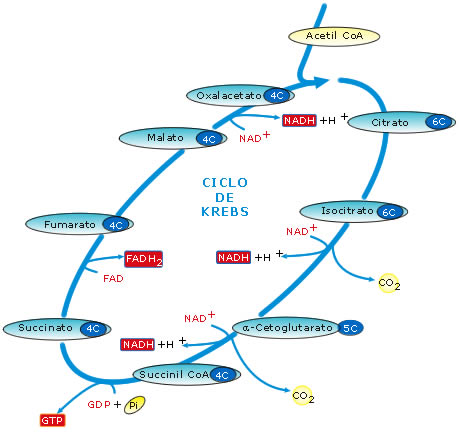
Para ver las reacciones del ciclo de Krebs con más detalle pincha aquí.
La cadena respiratoria
Los
transportadores de electrones NADH y FADH2, originados
fundamentalmente en el ciclo de Krebs, pero también en otros procesos catabólicos,
albergan el poder reductor que les confieren los electrones “energéticos”
que transportan.
Esa energía será liberada, poco a poco, a lo largo de la cadena respiratoria
que tiene lugar en las crestas y en la membrana mitocondrial interna. En
dicha membrana existen tres complejos enzimáticos transportadores de
electrones:
-
El complejo NADH deshidrogenasa
- El complejo citocromo b-c1<
- El complejo citocromo oxidasa.
Tanto
el NADH como el FADH2 ceden los electrones “energéticos” a la
cadena formada por esos tres transportadores y, a medida que pasan de un
transportador a otro, los electrones van liberando energía. Esa energía
(según la teoría quimiosmótica de Mitchell) permite el bombeo de
protones desde la matriz mitocondrial al espacio intermembranoso de la
mitocondria. De este modo se genera un gradiente electroquímico de protones,
con una concentración de protones mayor en el espacio intermembrana que en la
matriz.
La fuerza protón-motriz generada, impulsa los protones a través de las ATP
sintetasas presentes en la membrana mitocondrial interna, permitiendo la unión
del ADP a un grupo fosfato, con la consiguiente formación de ATP. El
conjunto de estos procesos, que culminan con la formación de ATP, constituyen
la fosforilación oxidativa.
Con fines prácticos, aunque no es del todo exacto, se considera que una molécula de NADH permite la formación de 3 moléculas de ATP, mientras que una de FADH2 sólo aportará 2 ATP.
Tanto los electrones como los protones, que han sido impulsados a lo largo de la
cadena respiratoria, deben unirse a un aceptor final. En la respiración
aerobia el aceptor último de electrones (y protones) es el O2, que
al unirse al H2, forma H2O como producto final.
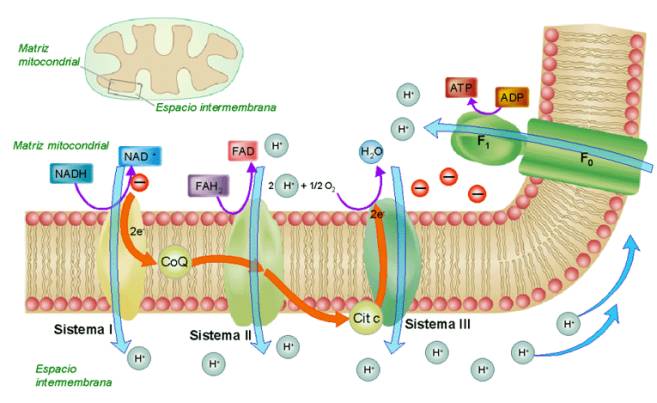
Transferencia
de electrones y fosforilación oxidativa
en la cadena respiratoria mitocondrial
Rendimiento energético del catabolismo aerobio de la glucosa
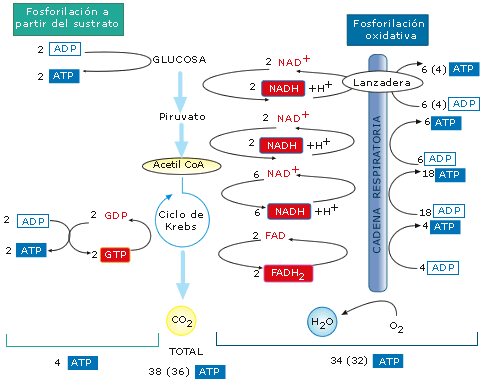
Balance
energético global de la respiración aerobia de
una
molécula de glucosa
B)
Vía anaerobia de las fermentaciones
El
piruvato formado en la glucólisis no siempre sigue la vía de la respiración
celular; en determinadas circunstancias puede pasar a la vía alternativa de las
fermentaciones.
Las fermentaciones genuinas son procesos anaerobios, realizados por
microorganismos que no toleran el oxígeno o por ciertas células animales o
vegetales cuando no disponen de suficiente oxígeno. Son poco rentables
desde el punto de vista energético, ya que la oxidación de la materia orgánica
es incompleta y se forma mucho menos ATP que en la respiración celular aerobia.
En general, únicamente 2 ATP por cada molécula de glucosa.
Dependiendo el producto final, se diferencian varios tipos de fermentaciones.
Las más importantes son: la fermentación alcohólica y la fermentación
láctica.
Fermentación alcohólica
En
la fermentación alcohólica o etílica, el piruvato se transforma en etanol
y se desprende CO2. La realizan, sobre todo, levaduras del género Saccharomyces
que tienen interés en la industria alimenticia por los productos residuales de
su metabolismo: el CO2 para esponjar la masa en la fabricación del
pan; y el etanol para producir diferentes bebidas alcohólicas (vino, sidra,
cerveza...).
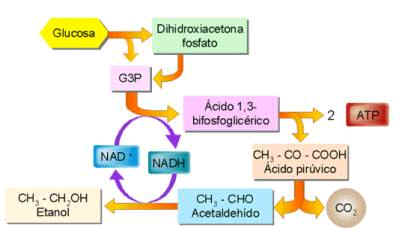
Fermentación láctica
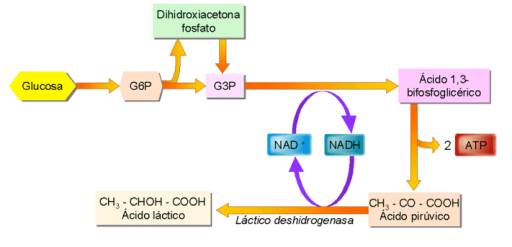
En ella el piruvato se transforma en lactato. La realizan diversas bacterias (Lactobacillus...) que fermentan la leche, y se utilizan para obtener derivados lácteos. Por otro lado, también la pueden llevar a cabo las células musculares cuando no reciben suficiente oxígeno. Así, cuando se realiza un esfuerzo intenso y prolongado, los músculos obtienen un poco de energía extra sin necesidad de oxígeno, recurriendo a la fermentación; pero las consecuencias de este proceso serán, posteriormente, las agujetas.
3.
EL CATABOLISMO DE LOS LÍPIDOS
Los ácidos grasos son los carburantes metabólicos más energéticos con que cuentan las células y en los animales desempeñan un papel de reserva energética fundamental debido a la facilidad que tienen para transformarse en grasas y almacenarse en los tejidos adiposos. La hidrólisis de las grasas, realizada por enzimas lipasas específicas, suministra importantes cantidades de ácidos grasos que son distribuidos por la sangre al resto de los tejidos, excepto al cerebro, que sólo utiliza glucosa como carburante metabólico.
Una vez en el citosol, los ácidos grasos para ser metabolizados se activan previamente en la membrana mitocondrial externa mediante el coenzima A, transformándose en acil-CoA, reacción que necesita un aporte energético del ATP.
El acil-CoA debe atravesar las membranas mitocondriales y, una vez en la matriz mitocondrial, sufre el ataque sucesivo de 4 sistemas enzimáticos que, de forma cíclica, van cortando fragmentos de 2 átomos de carbono a la cadena del acil-CoA y convirtiéndolos en moléculas de acetil-CoA, que continúan su proceso oxidativo en el ciclo de Krebs. La serie de 4 reacciones se repite hasta que el acil-CoA se ha oxidado totalmente a moléculas de acetil-CoA. Al conjunto de reacciones se le llama b-oxidación, debido a que el acil-CoA es oxidado siempre en su carbono b, es decir, el segundo que sigue al grupo ácido carboxílico. La serie de 4 reacciones que se repite es la siguiente:
§ Deshidrogenación, por medio de un FAD, del acil-CoA formándose un doble enlace. Se obtiene un deshidroacil-CoA.
§ Hidratación por medio de una molécula de agua que se fija en el doble enlace. Se obtiene un b-hidroxiacilCoA.
§ Oxidación del grupo alcohol a cetona reduciéndose un NAD. El producto resultante se llama b-cetoacil-CoA.
§ El compuesto anterior sufre una ruptura que recibe el nombre de tiolisis, liberándose acetil-CoA y formándose un acil-CoA con 2 carbonos menos. La tiolisis ocurre porque se fija un CoA en el carbono b.
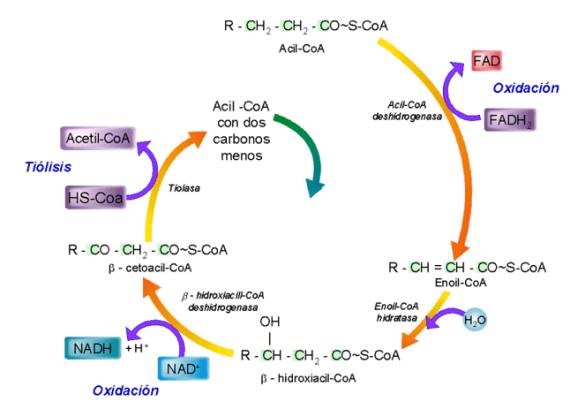
A partir del acil-CoA con dos carbonos menos obtenido, se continúa la b-oxidación repitiéndose las 4 reacciones hasta obtener otro acil-CoA, pero esta vez con 4 carbonos menos que el originario porque se forma un nuevo acetil-CoA. Y así sucesivamente hasta que el acil-CoA original se ha convertido en moléculas de acetil-CoA; en ese momento habrá completado la llamada hélice de Lynen.
El rendimiento energético de los ácidos grasos es muy superior al de los glúcidos. Como ejemplo pongamos el ácido palmítico. Este ácido tiene 16 carbonos, por tanto, para que se oxide completamente se necesitan siete vueltas del ciclo de la b-oxidación, de manera que en total se producen ocho moléculas de acetil-CoA. Teniendo en cuenta que cada molécula de acetil-CoA puede incorporarse al ciclo de Krebs, obtenemos lo que se representa en el siguiente cuadro:
|
8 Acetil-CoA
X 12 ATP/c. Krebs-cad. Resp. |
96 ATP |
|
7
FADH2 X 2 |
14 ATP |
|
7
NADH X 3 |
21 ATP |
|
TOTAL |
131 ATP |
Restando a esos 131 ATP obtenidos la molécula de ATP que se necesita para activar el ácido graso, resulta que un ácido palmítico puede proporcionar 130 ATP.
4.
EL CATABOLISMO DE LAS PROTEÍNAS
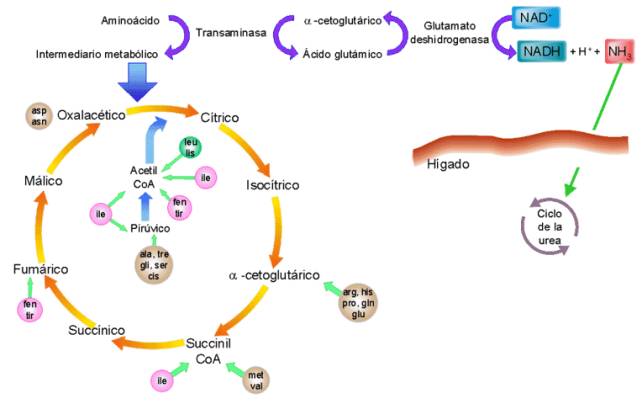
Las proteínas no son moléculas con una función energética fundamental. Pero en casos de necesidad, los aminoácidos producidos por la hidrólisis de las proteínas son degradados y sus derivados oxidados pueden entrar al ciclo de Krebs en algún punto determinado según el aminoácido.
Se distinguen tres mecanismos de degradación de los aminoácidos:
§ Transaminación, consiste en el traspaso del grupo amino de un aminoácido a una molécula llamada α-cetoácido, que lo acepta y se transforma en otro aminoácido. Las transaminaciones son catalizadas por las transaminasas.
§ Desaminación oxidativa, es la liberación directa de los grupos amino de los aminoácidos en forma de NH4+, y así se forman α-cetoácidos. Las desaminaciones son catalizadas por deshidrogenasas y producen coenzimas reducidos (NADH) que pueden entrar a la cadena respiratoria. El NH4+ formado se convierte en urea en la mayoría de vertebrados terrestres (ureotélicos), por medio del ciclo de la urea, y posteriormente es excretado. Otra forma de excreción, que utilizan organismos como aves y algunos reptiles (uricotélicos) es el ácido úrico. Los organismos acuáticos (amoniotélicos) suelen excretar el NH4+ directamente porque les es más fácil disolverlo en el medio.
§ Descarboxilación, es la pérdida del grupo carboxilo (-COOH) en forma de CO2, gracias al CoA. Previamente el aminoácido debe haber perdido el grupo amino. El producto puede incorporarse al ciclo de Krebs.