
GENÉTICA
MENDELIANA
1.
MENDEL: EL NACIMIENTO DE LA GENÉTICA
En 1865, el monje agustino austriaco Gregor Joham Mendel, abad del monasterio de Brünn (Chequia), formuló las leyes hereditarias que llevan su nombre, fruto de los estudios en su jardín con determinadas especies de guisantes (Pisum sativum).
Mendel trabajó sobre la transmisión
de los caracteres de las plantas a través de sucesivas generaciones, en lo que
hoy constituye el fundamento de la genética moderna.
Gracias a sus numerosos experimentos logró el enunciado de los principios conocidos más tarde como leyes de la herencia.
Su obra más
importante sobre la herencia fue publicada en 1866. Pero su trabajo no tuvo
trascendencia alguna en los siguientes treinta y cuatro años. No obtuvo el
debido reconocimiento hasta 1900, cuando tres investigadores (De Vries, Correns
y Tschermak), de manera más o menos independiente, llegaron a las mismas
conclusiones que él, descubrieron sus trabajos con gran sorpresa y le
reconocieron la prioridad, por eso publicaron sus conclusiones como meras
confirmaciones de las leyes de Mendel; y sólo a finales de la década de 1920 y
comienzos de 1930 se comprendió su verdadero alcance, en especial en lo que se
refiere a la teoría evolutiva.
Conviene aclarar que
Mendel, por ser pionero, carecía de los conocimientos actuales sobre la
presencia de pares de cromosomas en los seres vivos ni sobre su mecanismo de
transmisión, por lo que la interpretación de sus experimentos, que realizamos
más adelante, está basada en la posterior teoría cromosómica de la
herencia.
Mendel llegó a la conclusión de que los caracteres están
determinados por algo, a lo denominó factores o elementos hereditarios
(el equivalente a los actuales genes); los cuales se encuentran en parejas
antagónicas (hoy los llamamos genes alelos) y se transmiten con independencia
unos de otros (en este sentido tuvo suerte de no encontrarse con casos de
ligamiento y recombinación).
2.
TEORÍA CROMOSÓMICA DE LA HERENCIA
En la época en la que Mendel realizó sus investigaciones no se conocían
los genes ni el papel de la meiosis
en la herencia de los caracteres.
En 1902, W.S. Sutton en Estados Unidos y T. Boveri en Alemania,
propusieron la hipótesis de que los factores hereditarios de Mendel se
localizaban en los cromosomas, ya que creían que la separación de los cromosomas durante la meiosis
era la base para explicar las leyes de Mendel.
En 1911, T.H. Morgan, después de realizar numerosos experimentos con la
mosca de la fruta o del vinagre (Drosophila melanogaster) concluyó que
muchos caracteres se heredan juntos debido a que los genes (término
propuesto por W. Johannsen en 1909) que los codifican se encuentran juntos en un
mismo cromosoma, es decir, se hallan ligados. Nace así la teoría cromosómica
de la herencia, la cual ha tenido aportaciones posteriores, y hoy día puede
resumirse en los siguientes postulados:
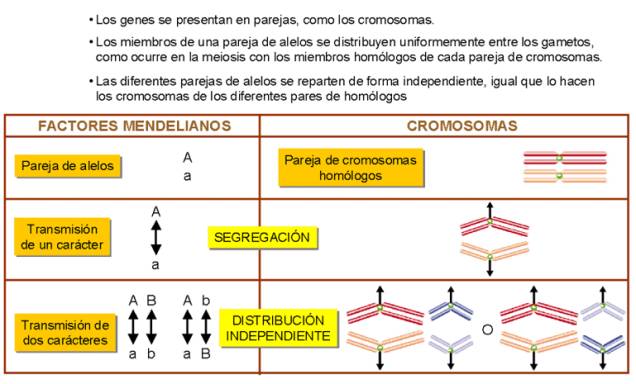
§
Los factores (genes) que
determinan los factores hereditarios del fenotipo se localizan en los cromosomas.
§
Cada gen ocupa un lugar
específico o locus (en plural es loci) dentro de un cromosoma concreto.
§
Los genes (o sus loci) se
encuentran dispuestos linealmente a lo largo de cada cromosoma.
§ Los genes alelos (o factores antagónicos) se encuentran en el mismo locus de la pareja de cromosomas homólogos, por lo que en los organismos diploides cada carácter está regido por una par de genes alelos.
Gen: Unidad hereditaria que controla cada carácter en los seres vivos.
Alelo: Cada una
de las alternativas que puede tener un gen de un carácter.
3. LEYES DE MENDEL: LAS REGLAS DE LA HERENCIA
Primera ley de Mendel
A
esta ley se le llama también ley de la uniformidad de los híbridos de la
primera generación (F1) o principio de dominancia, y dice que cuando se
cruzan dos variedades individuos de raza pura (homocigotos) para un determinado
carácter, todos los descendientes de la primera generación son iguales (heterozigotos
o híbridos).
Homocigoto o raza pura: Individuo que para un gen dado tiene en cada cromosoma homólogo el mismo tipo de alelo, por ejemplo, AA o aa.
Heterocigoto o híbrido: Individuo que para un gen dado tiene en cada cromosoma homólogo un alelo distinto, por ejemplo, Aa.
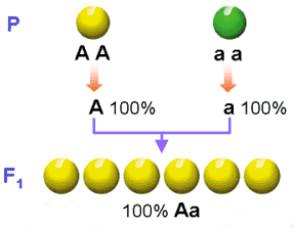 El experimento de Mendel: Mendel llegó a esta conclusión trabajando con una variedad pura de
plantas de guisantes que producía semillas amarillas y con una variedad que
producía semillas verdes. Al hacer un cruzamiento entre estas plantas, obtenía
siempre plantas que producían semillas amarillas.
El experimento de Mendel: Mendel llegó a esta conclusión trabajando con una variedad pura de
plantas de guisantes que producía semillas amarillas y con una variedad que
producía semillas verdes. Al hacer un cruzamiento entre estas plantas, obtenía
siempre plantas que producían semillas amarillas.
Interpretación
del experimento: el polen de
la planta progenitora aporta a la descendencia un factor hereditario (alelo)
para el color de la semilla, y el óvulo de la otra planta progenitora aporta
otro factor hereditario para el color de la semilla; de los dos factores,
solamente se manifiesta aquél que es dominante (A), mientras que el recesivo
(a) permanece oculto.
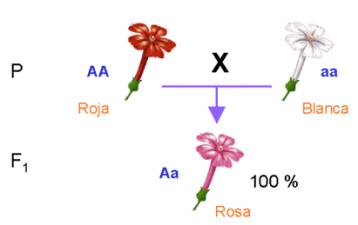 Otros casos para la primera ley: la primera ley de Mendel se cumple también para el caso en que un
determinado gen de lugar a una herencia intermedia, como es el caso del
color de las flores del "dondiego de noche" (Mirabilis jalapa).
Otros casos para la primera ley: la primera ley de Mendel se cumple también para el caso en que un
determinado gen de lugar a una herencia intermedia, como es el caso del
color de las flores del "dondiego de noche" (Mirabilis jalapa).
Al cruzar las plantas de la variedad de flor blanca con plantas de la
variedad de flor roja, se obtienen plantas de flores rosas.
La interpretación es la misma que en el caso anterior: cada individuo recibe un alelo del padre y un alelo de la madre, solamente varía la manera de expresarse los distintos alelos de los genes: no hay uno que domine sobre el otro y ambos se expresan son alelos codominantes.
Segunda ley de Mendel
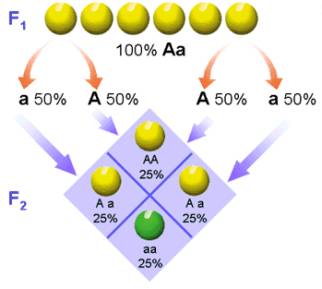
Del cruce obtuvo semillas amarillas y verdes en una proporción de 3:1.
Así pues, aunque el factor hereditario que determina la coloración
verde de las semillas parecía haber desaparecido en la primera generación
filial, vuelve a manifestarse en esta segunda generación.
Interpretación del experimento: los dos alelos distintos para el color de la semilla presentes en los individuos de la primera generación filial, no se han mezclado ni han desaparecido, simplemente se manifestaba sólo uno de los dos.
Cuando el individuo de fenotipo amarillo y genotipo Aa, forme los gametos, se separan los alelos, de tal forma que en cada gameto sólo habrá uno de los alelos y así puede explicarse los resultados obtenidos.
Genotipo: Es el conjunto de genes que contiene un organismo heredado de sus progenitores.
En organismos diploides, la mitad de los genes se heredan del padre y la otra mitad de la madre.
Fenotipo:
Es la manifestación externa del genotipo, es decir, la suma de los caracteres
observables en un individuo. El fenotipo es el resultado de la interacción
entre el genotipo y el ambiente. El ambiente de un gen lo
constituyen los otros genes, el citoplasma celular y el medio externo donde se
desarrolla el individuo.
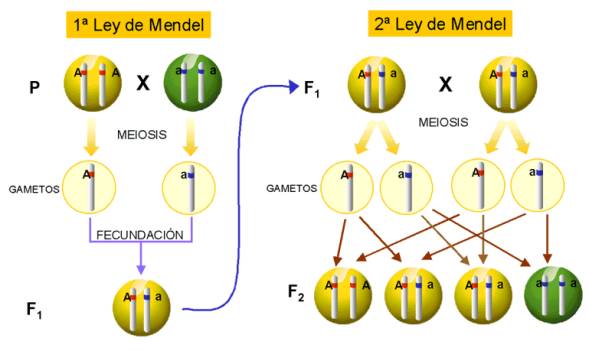
Intepretación
cromosómica de la primera y segunda ley de Mendel.
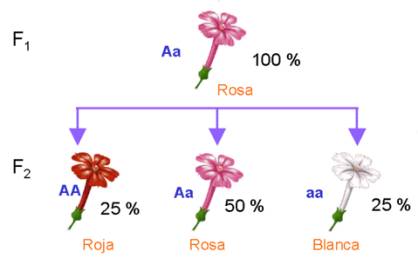
En el caso de la herencia intermedia, también se cumple la
segunda ley: si cruzamos entre sí dos plantas de flores rosas de la primera
generación filial, se obtienen plantas con flores blancas, rosas y rojas, en la
proporción 1:2:1.
También en este caso se manifiestan los alelos para el color rojo y blanco, que permanecieron ocultos en la primera generación filial.
Retrocruzamiento o
cruzamiento prueba
En el caso de
los genes que manifiestan herencia dominante, no existe ninguna diferencia
aparente entre los individuos heterocigóticos (Aa) y los homocigóticos (AA),
pues ambos individuos presentarían un fenotipo amarillo.
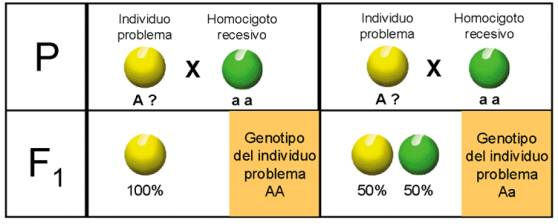
La prueba del retrocruzamiento, o simplemente cruzamiento prueba, sirve
para diferenciar el individuo homocigótico del heterocigótico.
Consiste en cruzar el fenotipo dominante con la variedad homocigota recesiva (aa). Si es homocigótico, toda la descendencia será igual, en este caso se cumple la primera ley de Mendel. Si es heterocigótico, en la descendencia volverá a aparecer el carácter recesivo en una proporción del 50 %.
Tercera
ley de Mendel
 Se conoce como ley de la independencia de caracteres, y hace
referencia al caso de que se contemplen dos caracteres distintos. Cada uno de
ellos se transmite siguiendo las leyes anteriores con independencia de la
presencia del otro carácter.
Se conoce como ley de la independencia de caracteres, y hace
referencia al caso de que se contemplen dos caracteres distintos. Cada uno de
ellos se transmite siguiendo las leyes anteriores con independencia de la
presencia del otro carácter.
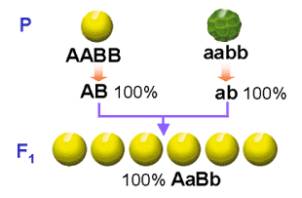
El experimento de Mendel: Mendel cruzó plantas de guisantes de semilla amarilla y lisa con plantas de semilla verde y rugosa (homocigóticas ambas para los dos caracteres).
Las semillas obtenidas en este cruzamiento eran todas amarillas y lisas,
cumpliéndose así la primera ley para cada uno de los caracteres considerados,
y revelándonos también que los alelos dominantes para esos caracteres son los
que determinan el color amarillo y la forma lisa.
Las plantas obtenidas y que constituyen la F1 son dihíbridas (AaBb).
Estas plantas de la F1 se cruzan entre sí.
Los gametos que formarán cada una de las plantas son:
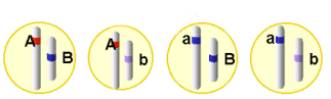
En
el esquema siguiente se ven los tipos de semillas que se originan en dicho cruce
y sus proporciones de aparición:
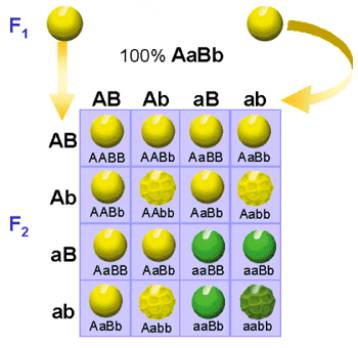
Se puede apreciar que los alelos de los distintos genes se transmiten con independencia unos de otros, ya que en la segunda generación filial F2 aparecen guisantes amarillos y rugosos y otros que son verdes y lisos, combinaciones que no se habían dado ni en la generación parental P, ni en la filial primera F1. Asímismo, los resultados obtenidos para cada uno de los caracteres considerados por separado, responden a la segunda ley.
Interpretación del experimento: los resultados de los experimentos de la tercera ley refuerzan el concepto de que los genes son independientes entre sí, que no se mezclan ni desaparecen generación trás generación.
Para esta interpretación fue providencial la elección de los caracteres, pues estos resultados no se cumplen siempre, sino solamente en el caso de que los dos caracteres a estudiar estén regulados por genes que se encuentran en distintos cromosomas. No se cumple cuando los genes considerados se encuentran en un mismo cromosoma (genes ligados).
4.
SERIES ALÉLICAS
Alelos múltiples
Hasta ahora, en todos los casos vistos, un gen tenía dos formas diferentes de expresarse (dos alelos); pero puede ocurrir que un mismo gen tenga múltiples formas de presentarse, en cuyo caso decimos que tenemos una serie de alelos múltiples. En estos casos, ha de establecerse la jerarquía de dominancia de unos alelos sobre otros.
Los alelos múltiples más conocidos dentro de la especie humana son los de los grupos sanguíneos AB0. Existen tres formas alélicas, representadas por los alelos "A", "B", "0". Los alelos A y B son codominantes entre sí y su característica principal es que cada uno de ellos codifica la síntesis de una proteína específica que se localiza en la superficie de los glóbulos rojos; son el antígeno A y el antígeno B. La forma alélica 0 es recesiva con respecto a los alelos A y B y a su vez no produce ningún antígeno.
La jerarquía de dominancia en este
caso es: A = B > 0
Genotipo |
AA,
A0 |
BB,
B0 |
AB |
00 |
|
Fenotipo |
A |
B |
AB |
0 |
5.
HERENCIA Y SEXO
La diferenciación sexual es la expresión fenotípica de un conjunto de factores genéticos que determinan que el individuo sea capaz de producir uno u otro tipo de células sexuales. Los individuos machos, o de sexo masculino, son los productores de espermatozoides; los individuos hembras, o de sexo femenino, son los productores de óvulos y los individuos hermafroditas son capaces de producir los dos tipos de gametos.
En algunas especies la determinación sexual es producida por un gen con varios alelos. Es el caso del pepinillo del diablo, en el que se dan tres alelos para la sexualidad:
|
ADAD
, ADA+ |
Masculino |
|
A+A+
, A+Ad |
Monoico |
|
AdAd
|
Femenino |
El caso más común de determinación sexual es aquél en el que los genes que determinan la sexualidad se reunen en unos cromosomas determinados que se llaman cromosomas sexuales.
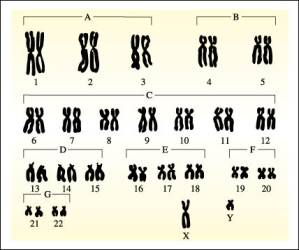
En todas las células de un individuo, excepto en los gametos, existen dos
series de cromosomas, que forman parejas de homólogos, es decir
representamos su dotación cromosómica por 2n.
Al agrupar estas parejas de homólogos
existe un par de cromosomas que es diferente según estemos estudiando una
hembra o un macho. Estos dos cromosomas, que pueden ser identificados por su
forma y tamaño como pertenecientes a uno de los dos sexos, se denominan cromosomas
sexuales, mientras que los restantes pares de cromosomas homólogos, que son
iguales en tamaño y forma para ambos sexos de una misma especie, se denominan autosomas.
La representación gráfica de todos los cromosomas de un individuo en cuanto al número, tamaño y forma constituye el cariotipo.
Los cromosomas sexuales se han denominado X e Y. En los mamíferos, las células de los individuos machos contienen un par XY y las células de las hembras por un par XX. En la especie humana, cuya dotación cromosómica es de 46 cromosomas, cada célula somatica contiene 22 pares de autosomas más un par XX si se trata de una mujer y 22 pares de autosomas y un par XY si se trata de un varón.
La determinación sexual queda marcada en el momento de la fecundación y viene fijada por el tipo de gametos que se unen. Las mujeres sólo produciran un tipo de óvulo con 22 autosomas y un cromosoma sexual X, mientras que los varones formaran dos tipos de espermatozoides, el 50 % portadores de un cromosoma X y el 50 % portadores de un cromosoma Y.
Al ser la fecundación producto del azar, un óvulo puede unirse a cualquiera de
los tipos de espermatozoides que se han producido, por lo que en la mitad de
los casos se formarán hembras y en otro 50% se formaran machos.
No siempre el individuo heterogamético es el
macho (como en el sistema XX/XY de los humanos). En aves, la hembra es heterogamética
y el macho es homogamético, para diferenciar este sistema se usan las letras Z
y W. En algunos insectos como los saltamontes el macho es heterogamético pero
con la particularidad de tener un solo cromosoma sexual.
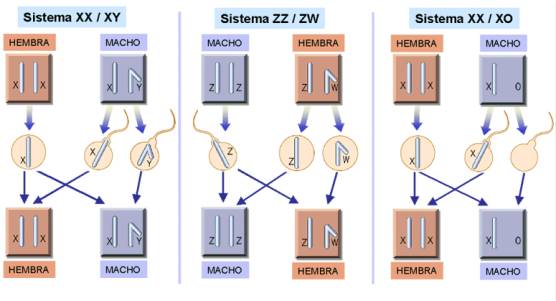
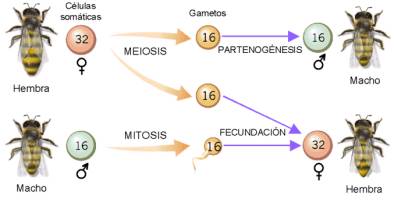
En himenópteros sociales (abejas, avispas...), el sexo viene determinado por la dotación cromosómica: los individuos diploides son hembras y los haploides son machos que se han desarrollado por partenogénesis (a partir de un óvulo no fecundado).
En muchas especies, la dotacón genética no determina totalmente el tipo de sexo, y son las condiciones ambientales las que realizan dicha determinación.
Muchas plantas superiores que
producen flores hermafroditas pueden masculinizar o feminizar sus flores cuando
se las trata con alguna fitohormona.
Las larvas de los gusanos marinos Bonellia viridis que nadan libremente
dan lugar a hembras, mientras que las larvas que se desarrollan en una hembra
dan lugar a machos.
En especies de ranas y sapos, los machos son hermafroditas en su fase juvenil y las hembras adultas pueden pasar a hermafroditas e incluso a machos.
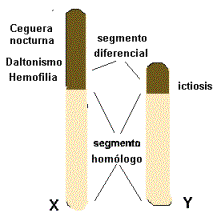 En
la especie humana los cromosomas X e Y presentan diferencias morfológicas
(el Y es más pequeño que el X) y tienen distinto contenido génico.
En
la especie humana los cromosomas X e Y presentan diferencias morfológicas
(el Y es más pequeño que el X) y tienen distinto contenido génico.
Están compuestos por un segmento homólogo donde se localizan genes que regulan los mismos caracteres y otro segmento diferencial, en este último se encuentran tanto los genes exclusivos del X , caracteres ginándricos, como los del cromosoma Y, caracteres holándricos.
Los caracteres cuyos genes se localizan en el segmento diferencial del cromosoma X, como daltonismo, hemofilia, ictiosis están ligados al sexo.
Consiste en la incapacidad de
distinguir determinados colores, especialmente el rojo y el verde. Es un
caracter regulado por un gen recesivo localizado en el segmento
diferencial del cromosoma X.
Los genotipos y fenotipos posibles son:
MUJER ♀ |
HOMBRE ♂ |
|
XDXD: visión normal |
XD Y: visión normal |
|
XDXd: normal/portadora |
Xd Y: daltónico |
|
XdXd: daltónica |
|
Se caracteriza por la incapacidad de coagular la sangre,
debido a la mutación de uno de los factores proteicos. Igual que en el
daltonismo, se trata de un carácter recesivo, y afecta fundamentalmente
a los varones ya que las posibles mujeres hemofílicas Xh Xh
no llegan a nacer, pues esta combinación homocigótica recesiva es letal
en el estado embrionario.
Los genotipos y fenotipos posibles son:
MUJER ♀ |
HOMBRE ♂ |
|
XHXH: normales |
XH Y: normal |
|
XHXh: normal/portadora |
Xh Y: hemofílico |
|
Xh Xh: hemofílica (no nace) |
|
Algunos genes situados en los autosomas, o en las zonas homólogas
de los cromosomas sexuales, se expresan de manera distinta según se
presenten en los machos o en las hembras. Generalmente este distinto
comportamiento se debe a la acción de las hormonas sexuales masculinas.
Como ejemplo de estos caracteres podemos citar en los hombres la calvicie, un
mechón de pelo blanco y la longitud del dedo índice.
Si llamamos "A" al gen de pelo normal y "a"
al gen de la calvicie: El gen "a" es dominante en
hombres y recesivo en mujeres.
Según esto podemos tener los siguientes genotipos y fenotipos para el pelo:
|
Genotipo |
Hombres |
Mujeres |
|
AA |
Normal |
Normal |
|
Aa |
Calvo |
Normal |
|
aa |
Calvo |
Calva |
6.
GENES LIGADOS
A medida que se avanzó en la investigación sobre la herencia, se fue descubriendo que existían pares de genes que no se heredaban en las proporciones que había encontrado Mendel. Es decir, no siempre se cumple la tercera ley.
Esta ley se cumple cuando los caracteres elegidos están regulados por genes situados en distintos cromosomas.
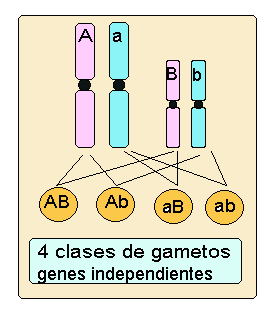
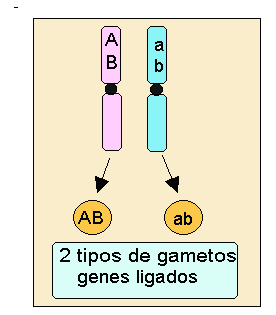
Como se aprecia en el esquema, los dos caracteres elegidos por Mendel color de la semilla "A" y forma de la semilla "B" se encuentran en distintos cromosomas y por lo tanto el individuo dihíbrido AaBb formará cuatro tipos de gametos (AB, Ab, aB, ab). En cambio si los genes que estamos estudiando se encuentran localizados en el mismo cromosoma, un individuo que tuviera el mismo genotipo dihíbrido AaBb sólo formará dos clases de gametos, en el caso concreto del esquema se formarán los gametos con las combinaciones: AB, ab.
En el caso concreto de las semillas de guisantes, como se ve en el esquema siguiente, en el que se ha utilizado la técnica del cruce de prueba, tendríamos estos resultados:
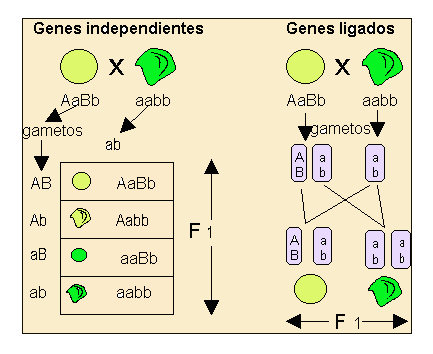
Hoy día sabemos que los cromosomas son portadores de un número elevado de genes, por lo tanto, cuando un cromosoma es heredado por un hijo también son heredados todos sus genes, y en este caso su comportamiento no sigue la Tercera Ley de Mendel, es decir, no se segregan independientemente.
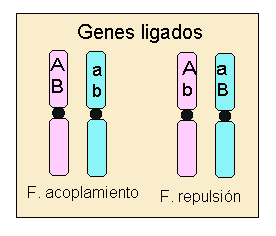
A los genes que están localizados en el mismo cromosoma se les llama
genes ligados.
Los genes ligados pueden encontrarse en fase de acoplamiento, alelos dominantes en el mismo cromosoma; o en fase de repulsión, alelo dominante de un carácter junto al recesivo para el otro carácter.
Entrecruzamiento
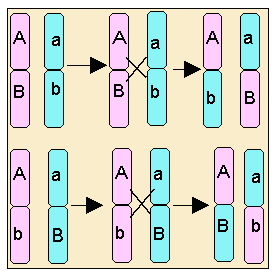 Sabemos
que durante la profase I de la meiosis se producen entrecruzamientos con una
determinada probabilidad. Si dos genes están alejados dentro del mismo
cromosoma, es posible que queden segregados porque la probabilidad de que se dé
un entrecruzamiento es más alta. En ese caso, aparecen
cromosomas recombinados.
Sabemos
que durante la profase I de la meiosis se producen entrecruzamientos con una
determinada probabilidad. Si dos genes están alejados dentro del mismo
cromosoma, es posible que queden segregados porque la probabilidad de que se dé
un entrecruzamiento es más alta. En ese caso, aparecen
cromosomas recombinados.
Como podemos ver aparecen de nuevo todas las combinaciones de gametos, como si la segregación fuera independiente, pero las frecuencias de aparición de los gametos recombinados son menores que las esperadas para una segregación independiente.
La frecuencia de aparición de recombinantes nos proporciona una idea de la colocación de los genes dentro de un cromosoma y con su ayuda se pueden trazar mapas cromosómicos.