|
© ASTRONET 2008 |
|
ASTRONET—Astrofísica en la Web |
|
La vida en nuestro planeta tiene asombrosas formas de adaptación, desarrollo, crecimiento. |

|
Todos los derechos reservados a este site. © ASTRONET 2007-2008 |
|
MODELOS SIMPLES DE POBLACIONES ANIMALES © ASTRONET 2008- Derechos reservados- Permitida su reproducción citando la fuente. Consideraremos qué cambios ocurren en una población de animales cuando un pequeño número de éstos irrumpe en un nicho ecológico apropiado. Existe la posibilidad de reconstruir una situación simple para el análisis, en condiciones de laboratorio. Los modelos experimentales se prestan a la expresión cuantitativa de un modo más simple que las investigaciones de campo. Supongamos entonces, un volumen limitado espacialmente que cumple con las condiciones físicas de un nicho ecológico que es apto para alguna especie dada. Supongamos también que existe una circulación apropiada de nutriente para este sistema. Dos situaciones de equilibrio son posibles en dicho espacio: 1.– Equilibrio inestable: ocurre cuando los organismos están excluidos de la incorporación al espacio en cuestión. 2.- Equilibrio originado en un receptáculo que esté saturado de animales tal que no pueda incrementarse su número. Un espacio de estas características puede ser representado mediante el gráfico de Verhulst logístico. A medida que la densidad de animales del ambiente aumenta, va disminuyendo la tasa de aumento por animal en proporción a la densidad, hasta que finalmente, el aumento cesa por completo. Se realiza una serie de suposiciones, tendientes a simplificar el modelo con objeto a su estudio: i– Todos los animales de la población poseen la misma posibilidad de morir, reproducirse, comer, o ser comidos. Es decir, tendrán idénticas propiedades ecológicas. Llamamos N al número de animales existentes en la población. Este número puede medirse en unidades de masa o en cantidad de individuos. ii– Todos los animales de la población responden de manera instantánea (sin retardo alguno) a las alteraciones del medio. El cambio en la población puede representarse como: dN/dt = f(N) iii– Se define como capacidad de carga K al límite superior, para una dada población y un dado ambiente, al cual crecerá el tamaño de la población. De esto se concluye que dado un medio, un nutriente, y una población única, en condiciones de laboratorio se puede realizar un modelo de estudio simple, que tomará su forma más realista y adecuada a medida que cada parámetro involucrado sea considerado en sí mismo como función de las definiciones más complejas del sistema. ______________________________________________________________________________ |

|
Modelos simples de poblaciones animales El moho mucilaginoso La filotaxis Crecimiento de bacterias en un quemostato |
|
EL MOHO MUCILAGINOSO © ASTRONET 2008- Derechos reservados- Permitida su reproducción citando la fuente. Un grupo de organismos que se conocen como mohos mucilaginosos celulares (Dictyostelium discoideum), posee un sistema de comunicación celular. Al llegar a una fase dada de su ciclo vital, se asemeja a un conjunto de pequeñas células libres, ameboides, que se dividen, crecen y alimentan, hasta que su reserva de nutrientes (en este caso, bacterias) finalmente se agota. Al llegar a esta nueva etapa, las células del moho alteran · su forma · su comportamiento. Adquieren un aspecto alargado y empiezan un trabajo de migración hacia el centro del grupo de ameboides. Posteriormente, se apilan, formando una masa multicelular que migra del lugar sin nutrientes hasta que se asienta en otro, formando un montículo. Crece entonces un pie largo y delgado en cuyo vértice se forma un cuerpo fructífero, que madura y estalla, liberando un nuevo conjunto de células ameboides. El ciclo vuelve a iniciarse. La sustancia química que se transmite entre las células para realizar este proceso conjunto se llama AMP cíclico (cAMP), y se trata de monofosfato de adenosina, derivado de la molécula de ATP. |
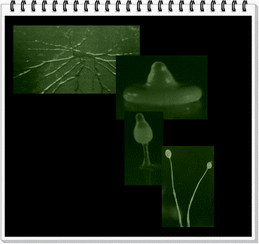


|
La forma en que la hoja se une al tallo es muy variable. En casi todas las dicotiledóneas, el punto de unión es la base del peciolo, mientras que éste suele faltar en las hojas de las monocotiledóneas, cuya base se transforma en una vaina ancha y plana que envuelve el tallo. Algunas hojas pecioladas forman en el punto en que se unen al tallo una estructura semejante a una hoja o escama llamada estípula, como se observa en las Rosáceas. Se conocen dos formas básicas de filotaxis: alterna, con las hojas dispuestas alrededor del tallo en espiral; y opuesta, con las hojas unidas al tallo por parejas a la misma altura; si las parejas sucesivas se disponen formando ángulo recto, como en la menta, se dice que las hojas están decusadas; y si de un mismo punto del tallo brotan más de dos hojas, se habla de hojas verticiladas.
|
|
LA FILOTAXIS |
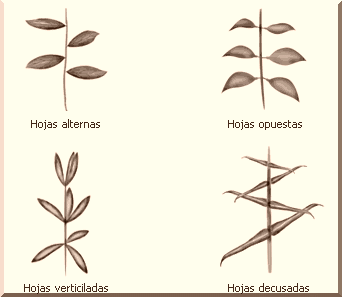
|
CRECIMIENTO DE BACTERIAS EN UN QUEMOSTATO © ASTRONET 2008- Derechos reservados- Permitida su reproducción citando la fuente. Un sistema compuesto por un reservorio de nutriente unido a una cámara de cultivo bacterial es comúnmente designado como quemostato. Se utiliza en investigaciones de laboratorio de modelos poblacionales para mantener niveles de población convenientes al estudio. En 1880 se inició el conocimiento científico de la inmunidad frente a las bacterias: Pasteur descubrió que el Bacillus anthracis cultivado a una temperatura entre 42 y 43 °C pierde toda su virulencia tras varias generaciones, y más tarde se descubrió que los animales inoculados con estas bacterias debilitadas eran resistentes a la infección. Desde esa fecha se puede decir que nació la prevención, modificación y tratamiento de las enfermedades mediante la inmunización, uno de los avances más importantes de la medicina actual El sistema se diseña tal que la tasa de flujo que pasa del reservorio de nutriente a la cámara que contiene las bacterias no sea tan grande que barra todo el cultivo, e decir, que éste sea lavado y eliminado. La tasa de reabastecimiento del nutriente es suficientemente rápida como para que el cultivo siga creciendo. Se eligen entonces los parámetros: stock de concentración de nutriente tasa de entrada y salida de flujo tamaño de la cámara de crecimiento. |