Protokoll
zum Biochemischen Grundkurs 1998 4.11.98
Versuchstag 3: Puffer, Hämoglobin, Cytochrom c
von Dirk Hockemeyer
und Thomas Sandmann
Betreuer: Stefan Broer
Eingangsstempel:
Auswertung zum Versuchstag 3
1.Teil: Puffer
Aufgabe 1)
Diagramm A: Einstellung des Gleichgewichts
beim Begasen einer 25mM Natriumhydrogencarbonatlösung mit 5% CO2
in Stickstoff.
Diagramm B: Begasen einer Lösung der
gleichen Zusammensetzung mit Atemluft.
Diagramm C: Zugabe von 1 ml 0,1M Essigsäurelösung
zu einer Lösung der oben genannten Zusammensetzung.
Aufgabe 2)
Berechnet werden soll der pKsë-Wert des
offenen Puffersystems Kohlensäure-Hydrogencarbonat.
Dabei kann von Gleichung (13) ausgegangen werden:
Log (pCO2) = -pH + log[HCO3-]
+ pKsë ó log kH
Dafür muß die Gleichgewichtskonzentration
des Hydrogencarbonat-Ions bekannt sein.
Um diese zu berechnen, wird zunächst über
den Anfangs-pH-Wert 8,59 der Lösung die Protonenkonzentration berechnet.
[H+] = 2,57 . 10-9
mol/l
Geht man vom pH-Wert von 7 des destillierten Wassers
aus, sind einige Protonen durch die Reaktion mit Hydrogencarbonat zu Kohlensäure
verlorengegangen.
Es ergibt sich eine Ausgangskonzentration an Hydrogencarbonationen
von
[HCO3-] = 24,9999 mol/l.
Nach Einstellung des Gleichgewichts erhält man
einen pH-Wert von 7,51. Das entspricht einer Protonenkonzentration von
[H+] = 3,09 . 10-8
mol/l.
Es sind Protonen durch die Reaktion von CO2
und Wasser zu Kohlensäure und deren Dissoziation hinzugekommen. Dabei
wurde pro Proton ein Hydrogencarbonat-Ion gebildet, so daß die Gleichgewichtskonzentration
berechnet werden kann:
[HCO3-]GG= [HCO3-]Anfang
+ D [H+]
= 24,9999 . 10-3 mol/l.
Die eingesetzte Konzentration von 25mmol/l hat sich
kaum geändert. Deshalb kann ohne größeren Fehler mit der
Ausgangskonzentration gerechnet werden.
Mit Gleichung (13) ergibt sich sich ein pKsë-Wert
von 6,16.
Aufgabe 3)
Wie bei Aufgabe 2 muß zur Bestimmung des CO2-Gehaltes
wieder die Gleichgewichtskonzentration des Hydrogencarbonat-Ions bestimmt
werden. Auch hier kann mit 25mmol/l gerechnet werden, da wir uns in der
gleichen Ausgangslösung im gleichen pH-Bereich bewegen.
Bei angestrengtem Ausatmen konnte als niedrigster
pH-Wert 7,53 erreicht werden. Deshalb wird dieser Wert zur Rechnung benutzt.
Mit Gleichung (12) ergibt sich: pCO2=
35,43 torr
Aufgabe 4)
-
Bei Zugabe von 1 ml 0,1 M Essigsäure zum System
ohne Gaseinleitung (geschlossenes System) ändert sich der pH-Wert
um 0,49 Einheiten. Wird dann der Gasfluß eingeschaltet (offenes System),
nähert er sich wieder dem Ausgangswert an; die nderung ist nur noch
0,07 Einheiten groß.
Damit ergibt sich nach Gleichung (1) :
P = D
H+/D pH
die gemessene Pufferkapazität (siehe Tabelle).
-
Die theoretische Pufferkapazität ist als reziproke
Steigung der Neutralisationskurve am Punkt der Säure- bzw. Basenzugabe
derfiniert, damit ergibt sich nach Gleichung 4:
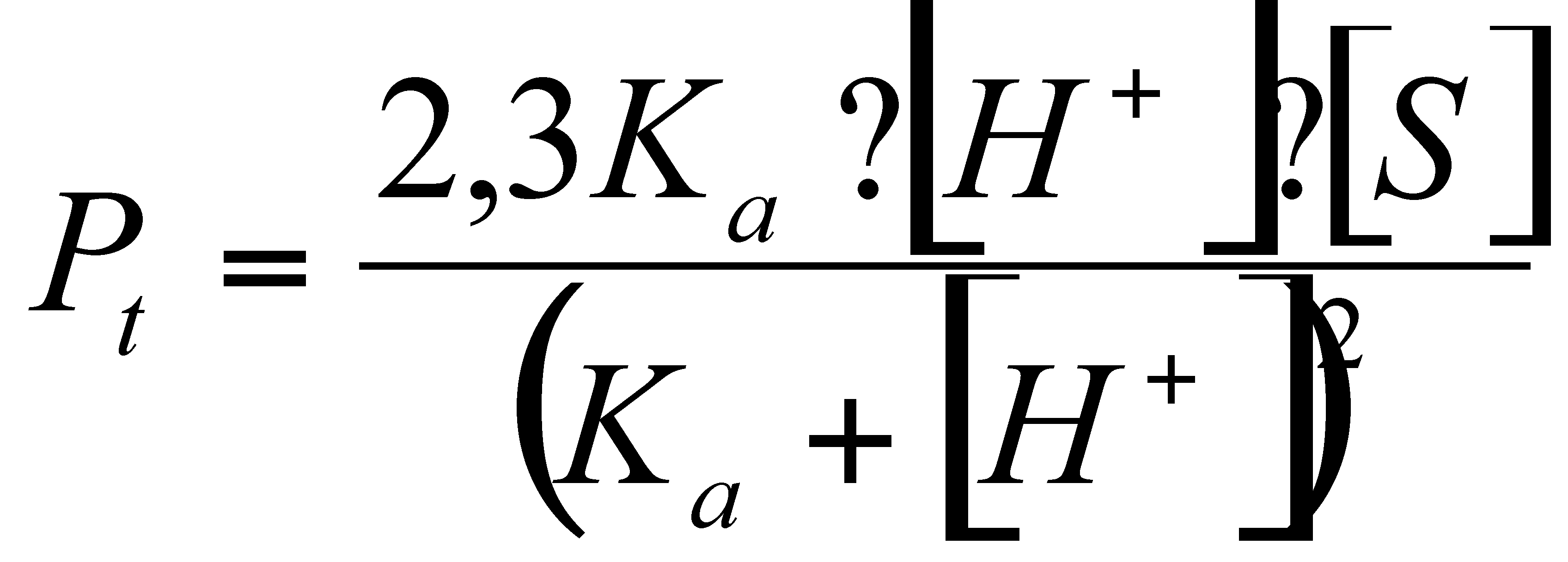
Dabei ist Ka in unserem Fall Ks
= 6,1 und [H+] die Protonenkonzentration im Gleichgewicht vor
Säurezugabe.
-
Die erwartete Pufferkapazität ergibt sich aus der
Henderson-Hasselbalch-Gleichung, wenn man den pH-Wert errechnet, der sich
nach der Zugabe von 0,1 mmol Protonen einstellt. Da dabei diese Menge an
Hydrogencarbonat verbraucht wird, wird von den 6,25.10-4
mol (entspricht der Ausgangsstoffmenge der 25 mM Lösung in 25 ml)
0,1 mmol abgezogen. Dieselbe Menge wird zur gelösten CO2-Menge
addiert.
Man erhält folgende Gleichung:
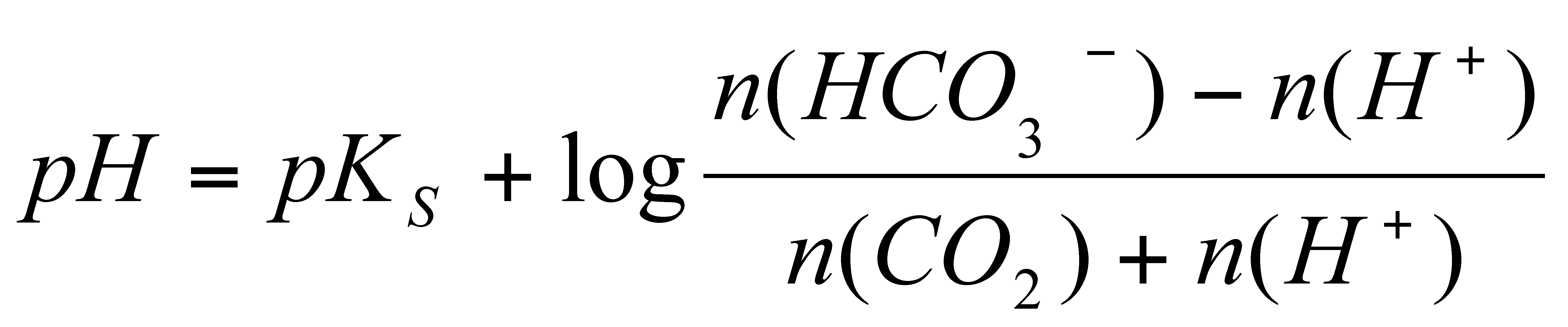
Damit ergibt sich in unserem Fall mit einer gelösten
CO2-Menge von
2,8219 . 10 -5 mol folgende
Gleichung:
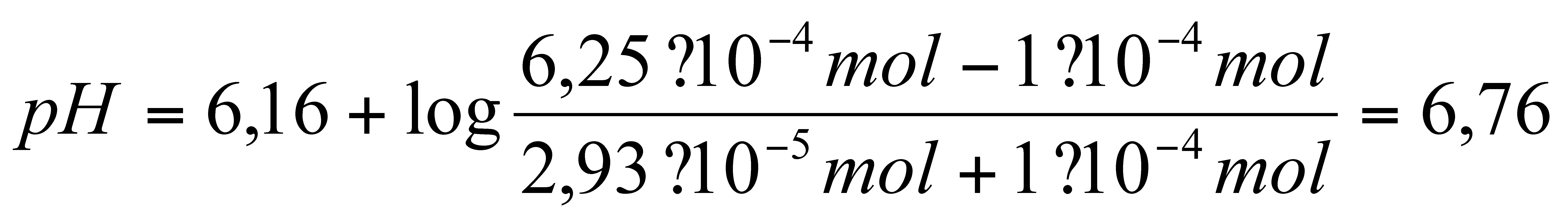
für das geschlossene System bzw.
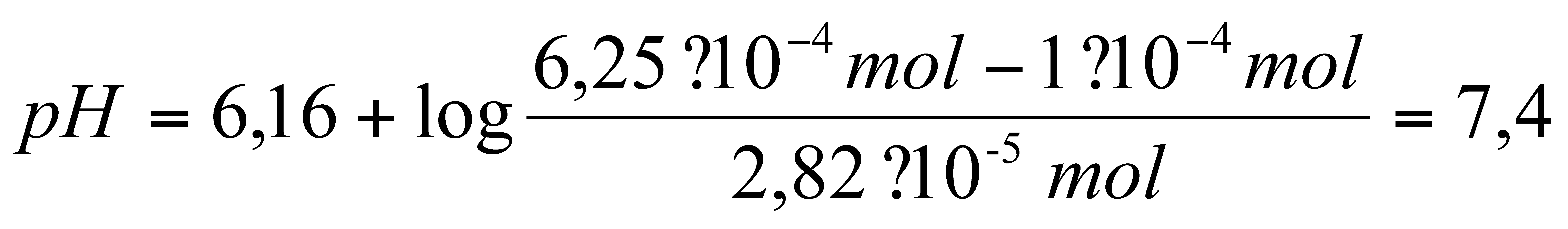
für das offene System.
Damit erhalten wir eine pH-Differenz vom Gleichgewichtswert
7,5 von
Geschlossenes System: 0,74
Offenes System:0,1.
Nach Gleichung (1) können nun die Pufferkapazitäten
bestimmt werden (siehe Tabelle).
-
Die praktische Pufferkapazität ist die Menge an
H+ / OH- pro Liter, die zugegeben werden muß,
bis sich der pH-Wert um eine Einheit ändert. Um sie zu bestimmen wird
die Henderson-Hasselbalch-Gleichung in der in 3. Benutzten Form nach [H+]
umgestellt.
Offenes System:
geschlossene System:
| |
D
H+/D pH
gemessen im Experiment |
D
H+/D pH
(erwartet)
(HH-Gleichung) |
D
H+/(D pH=1)
(praktisch)
(HH-Gl.) |
dH+/dpH (theoretisch)
(Gl. 4) |
| Geschlossen |
7,85.10-3
mol/l |
5,2.10-3
mol/l |
6,75.10-3
mol/l |
2,1 .10-3
mol/l |
| Offen |
0,055 mol/l |
0,038 mol/l |
0,026 mol/l |
------------------- |
Aufgabe 5)
Die erwartete und die gemessene Pufferkapazität
sollten natürlich schon übereinstimmen, allerdings können
bei der Messung verschiedene Fehlerquellen aufgetreten sein:
-
Falsches Eichen der Meßelektrode
-
Pipettierfehler und daraus resultierende Konzentrationsabweichungen
(besonders wahrscheinlich, da im Vergleich zu den anderen Gruppen ein sehr
hoher pH-Wert, sowohl als Ausgangswert als auch während der Messung,
aufgetreten ist.)
-
Nach Zugabe der Säure zu früh gemessen: kein
Gleichgewicht
-
Kein wirklich geschlossenes System
-
Schwankungen in der CO2-Menge durch Auf-
und Zudrehen der verschiedenen Hähne im Versuchssaal
-
Verunreinigungen im destillierten Wasser (z.B. durch
CO2)
Wie bei allen Messungen wir nicht ein reines Kohlensäure/Bicarbonat-GG
betrachtet, sondern es besteht auch noch die Gleichgewichtseinstellung
mit dem gasförmigen CO2. Deshalb müssen beide GG beachtet
werden und der Gesamt-pKs-Wert ist 6,1.
Auswertung zum Versuchstag 3
3.Teil: Cytochrom c
Aufgabe 1)
Diskussion des Kurvenverlaufs:
Erklärung der Linien:
-
Bis A: Cytochrom c-Lösung und Wasser, quilibierungsphase
-
A: Zugabe von 0,06 ml frischer Ascorbinsäurelösung
Das Cytochrom wird durch die Ascorbinsäure
reduziert, die reduzierte Form zeigt bei 550 nm eine starke Absorption,
die Extinktion steigt deshalb stark an.
-
B: Zugabe von 0,04 ml Cytochromoxidase-Lösung
Das Enzym oxidiert das Eisen der Hämgruppe
im Cytochrom c wieder, die Extinktion nimmer stark ab, da die oxidierte
Form wiederhergestellt wird.
-
C: Zugabe von 0,04 ml Ascorbinsäurelsg.
Erneut wird das Cytochrom c reduziert, da aber
auch das oxidierende Enzym in der Lösung vorliegt, wirkt sich die
Zugabe nicht so stark aus, wie bei A. Stattdessen erhalten wir ein Fließgleichgewicht,
da reduziertes Cytochrom c von der Cytochromoxidase wieder oxidiert wird.
Durch die hohe Aktivität des Enzyms liegt das Gleichgewicht der Redox-Reaktion
weit auf der oxidierten Seite: nur wenig reduziertes/absorbierendes Cytochrom
c ist vorhanden.
-
D: Erneute Zugabe von 0,04 ml Ascorbinsäurelsg.
Damit sollte das Fließgleichgewicht im Extinktionsdiagramm
deutlicher gemacht werden. Dies konnte leider nicht erreicht werden. Da
frische Ascorbinsäure benutzt wurde, kann aber auf diesem Wege ausgeschlossen
werden, daß die reduzierende Wirkung des Ascorbates aufgrund zu langer
Lagerung schon hätte verbraucht sein können.
-
E: Zugabe von 0,04 ml KCN-Lsg.
Die Cyanid-Ionen vergiften die die Cytochromoxidase,
damit ist das Fleißgleichgewicht aufgehoben und die reduzierte Form
wird nicht sofort wieder verbraucht.
-
F: Zugabe von 0,04 ml Kaliumhexacyanoferrat.
Dieses starke Oxidationsmittel setzt das reduzierte
Cytochrom c wieder in die oxidierte Form um und oxidiert die Ascorbinsäure
Es werden zweimal verdünnte Lösungen zugegeben, aber beide Male
reicht die Menge nicht aus, um alle reduzierten Stoffe umzusetzen. Erst
bei Zugabe von konzentrierter Kalium-Hexacyanoferrat-Lsg. (G) bleibt die
Extinktion auf niedrigem Niveau; das Cytochrom c wird nicht wieder reduziert.
Aufgabe 2)
Am Ende des Versuches wird Kaliumhexacyanoferrat
im Überschuß zugegeben. Alle reduzierten Formen werden daraufhin
oxidiert. Es ist davon auszugehen, daß nun alles Cytochrom c oxidiert
worden ist. Hier wird eine Extinktion von 0,384 gemessen.
Vollständig reduziert ist das Cytochrom nicht
nach der ersten Zugabe von Ascorbinsäurelösung, obwohl der Peak
hier am höchsten zu sein scheint. Berücksichtigt man aber die
Verdünnung (siehe unten), indem man die Extinktion mit dem Volumen
multipliziert, erhält man eine Extinktion in Abhängigkeit von
der Stoffmenge, die an Punkt F (vor der Zugabe von Hexacyanoferrat) am
höchsten ist. Allerdings steigt die Extinktion noch ganz leicht an,
so daß noch nicht die gesamte Menge umgesetzt worden sein kann. Nichtsdestotrotz
werden wir diesen höchsten stoffmengenabhängigen Extinktionswert
von 0, zur Berechnung heranziehen.
Für die Extinktion gilt die Gleichung:
E = e
.
c . d
Um die Extinktionsänderung beim Übergang
von der reduzierten zur oxidierten Form zu verfolgen, werden die oben genannte
Extinktionswerte benutzt. Durch die Zugabe von Lösungen wird allerdings
das Volumen vergrößert, damit verringert sich die Cytochrom-c-Konzentration.
Insgesamt werden bis zum Ende der Aufzeichnung 0,34 ml Lösung hinzugefügt.
Anfangsvolumen: 1 ml -> Cytochrom-c-Konzentration:
0,05 m mol/ml
Volumen vor der Hexacynaoferratzugabe: 1,22 ml
-> Cytochrom-c-Konzentration: 0,041 m
mol/ml
Endvolumen: 1,34 ml -> Cytochrom-c-Konzentration: 0,037
m mol/ml
Im Endvolumen soll, nach den obigen Überlegungen,
nur noch oxidierte Cytochrom c vorliegen. Vor der ersten Hexacyanoferrat-Zugabe
soll nur reduzierte Cytochrom c vorliegen.
D (E.V)= 0,000387
[Volumen in Liter]
Mit der Bildung der Differenz können wir das
"Grundrauschen", also die nicht von der reduzierten Form des Cytochroms
stammende Absorption herausrechnen.
Jetzt gilt: D
(E.V)= e
. n . d = 0,000387
Eingesetzt wurden 50.10-6 nmol
an Cytochrom c. Damit ist e
= 7,74 mM . cm-1. Die große Abweichung vom
Literaturwert 17,85 mM . cm-1 ist damit zu erklären,
daß die Annahme, bei 550 nm absorbiere nur die reduzierte Form des
Cytochroms c nicht richtig ist. Sie absorbiert aber deutlich stärker.
Außerdem haben wir ein relativ komplexes Gemisch erzeugt (Protein,
Ascorbinsäure,...), so daß nicht auszuschließen ist, daß
die Extinktion noch durch andere Einflüsse verfälscht wird. Zudem
treten noch die üblichen Fehlerquellen (falsches Ansetzen der Lösungen,
Pipettierfehler, etc.) auf den Plan, ganz zu schweigen vom oben genannten
Fakt, daß wir nicht davon ausgehen können, mit 0,739 schon die
maximale Extinktion in die Rechnung einbezogen zu haben.
Aufgabe 3)
In vivo kann die Ascorbinsäure das Cytochrom
nur oxidieren, wenn ein lipophiler Redoxvermittler hinzugefügt wird.
Das könnte ein Hinweis darauf sein, daß die Ascorbinsäure
die Hämgruppe des Cytochroms nicht erreichen kann, z.B. weil diese
auf die innere Lipiddoppelschicht des Mitochondriums gerichtet ist.