Glykolyse
Aufgabe 1:
Phosphatbestimmung:
In diesem Versuch wurde die Konzentration einer Fructose-1,6-bisphosphat-Lösung bestimmt.
Dazu wird das in der Lösung enthaltene Phosphat über die
Extinktion bei 820 nm eines Molybdenkomplexes bestimmt.Für eine Eichgrade
wurden die Extinktionen von bekannten "Standard"- Phosphatkonzentrationen
bestimmt.
| Standardlösung in µl in der Küvette | Stoffmenge in der Küvette in nmol | Konzentration in der Küvette in nmol.ml-1 | Extinktion |
| 0 | 0 | 0 | 0 |
| 20 | 10 | 8,33 | 0,054 |
| 50 | 25 | 20,83 | 0,234 |
| 80 | 40 | 33,33 | 0,454 |
| 110 | 55 | 45,83 | 0,695 |
| 140 | 70 | 58,3 | 1,093 |
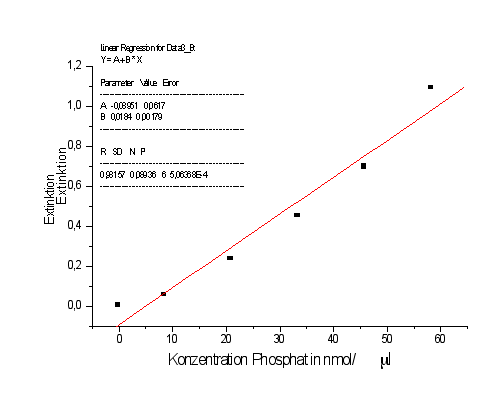
Trägt man nun die Extinktion gegen die Phosphatkonzentration auf, erhält man eine Gerade.
Über die Gradengleichung läßt sich nun den Extinktionswerten der Probelösung eine Konzentration zuordnen.
Geradengleichung:
Y=A+BX
Mit A=-0,08951 und B=0,0181
| Probennummer | Probenlösung in der Küvette in µl | Extinktion | Konzentration der Probenlösung in der Küvette in nmol.ml-1 |
| 1 | 30 | 0,119 | 11,52 |
| 2 | 50 | 0,293 | 21,13 |
| 3 | 70 | 0,474 | 31,13 |
Aus der Konzentration läßt sich nun die
Stoffmenge an Phosphat in der Küvette berechnen . (Volumen 1,2 ml)
| Probennummer | Stoffmenge in der Küvette nmol |
| 1 | 13,82 |
| 2 | 25,36 |
| 3 | 37,356 |
Diese Stoffmenge war in 30 µl (Probe 1), 50µl (Probe 2) bzw. 70µl (Probe 3) enthalten.
Daraus ergibt sich die Konzentration der Lösung,
in der das Phosphat in HCl gelöst wurde und damit die Stoffmenge an
Phosphat in dieser Lösung:
| Probennummer | Konzentration des Phosphates in der HCl-Lösung in nmol.µl-1 | Stoffmenge des gesamten Phosphates in der in HCl aufgenommen Lösung in nmol |
| 1 | 0,461 | 921,6 |
| 2 | 0,507 | 1014,24 |
| 3 | 0,534 | 1067,4 |
Daraus läßt sich nun die Fructose-1,6-bisphosphatkonzentration der Lösung berechnen.
Zu beachten ist, daß die Phosphatkonzentration
nach dem Aufschluß doppelt so groß ist, wie die Konzentration
des Fructose-1,6-bisphosphates der zu untersuchenden Lösung. Von dieser
Lösung wurden 5 µl eingesetzt.à
| Probennummer | Konzentration der Fructose 1,6 bisphosphat-Lösung in µmol.ml-1 |
| 1 | 92,16 |
| 2 | 101,42 |
| 3 | 106,74 |
Der Mittelwert dieser Ergebnisse liefert eine Konzentration für die Fructose 1,6 bisphosphat-Lösung von : 100,10 µmol/ml
Der angegebene Wert war 100µmol/ml à Abweichung 0,1%
Aufgabe 3
In diesem Versuch wird die Entkopplung, Vergiftung und die Entgiftung der Glycerinaldehyd-3phosphatdehydrogenase untersucht.
Durch die Zugabe von Arsenat in das Reaktionsgemisch
wird in der Reaktion der Glycerinaldehyd-3-phosphatdehydrogenase nicht
das energiereiche Phosphorsäureanhydrid sondern das Arsensäureanhydrid
gebildet. Dieses Anhydrid hydrolysiert dann unter der Bildung von 3-Phosphoglycerat
spontan und schnell. Diese Reaktion ist dann nicht mehr zu einem Energiegewinn
durch Phosphorylierung von ADP befähigt. Findet diese Reaktion in
der Glykolyse statt, wird keine Energie durch die Glykolyse gewonnen. Durch
diese stark exergonische Hydrolyse liegt das Gleichgewicht der Reaktion
weit auf der Seite des 3-Phosphoglycerates.
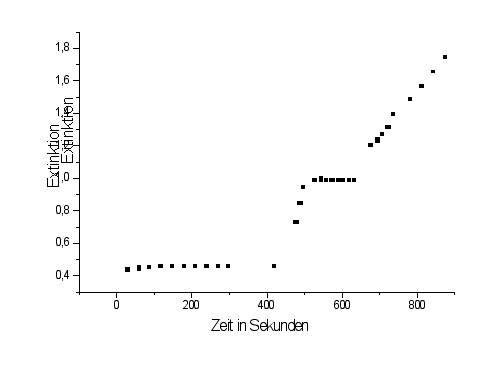
P-Chlormercuribenzoat reagiert mit Sulfhydrylgruppen. Diese werde oxidiert. Die Glycerinaldehyd-3-phosphatdehydrogenase enthält in ihrem aktiven Zentrum eine solche Gruppe, die das Glycerinaldehyd bindet. Wird diese Sulfhydrylgruppen oxidiert, kann keine Bindung erfolgen. In dem Diagramm erkennt erkennt man, daß nach der Zugabe von p-Chlormercuribenzoat kein weiteres NADH gebildet wird. Das Enzym ist vergiftet.
Durch die Zugabe von einem Überschuß von Mercoptoethanlol kann der Effekt des p-Chlormercuribenzoats rückgängig gemacht werden. Die Schwefelgruppe des Enzyms wird wieder zu einer Sulfhydrylgruppen reduziert und das p-Chlormercuribenzoats vom Enzym verdrängt. Nach der Zugabe von Mercaptoethanol erkennt man im Diagramm, daß wieder NADH gebildet wird. Das Enzym ist wieder aktiv. Es wurde von dem Mercaptoethanol entgiftet.
Versuch: Gekoppeltes Gleichgewicht von Aldolase-
und Gycerinaldehy-3-Phosphatdehydrogenase-Reaktion und Triosephosphatisomerase-Reaktion:
In diesem Versuch sollte die "biochemische" Gleichgewichtkonstate der Glycerinaldehyd 3-phosphatdehydrogenase und der Triosephosphatisomerase ( Gykolyseenzyme) bestimmt werden.
Die Reaktionen wurden photometrisch bei 405 nm über die Bildungsgeschwindigkeit von NADH verfolgt.
Aus der Differenz der Extinktion im Gleichgewicht nach der Phosphatzugabe und der Gleichgewichtsextinktion ohne Phosphat, läßt sich die Konzentration an gebildetem NADH bestimmen.
Die Extinktionswerte ergeben folgendes Diagramm: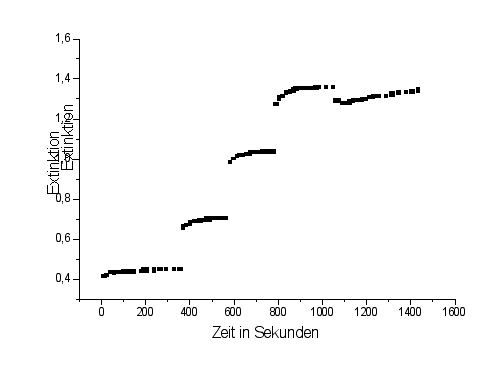
| Phosphatzugabe | Extinktion nach der Phosphatzugabe ó Extinktion ohne Phosphatzugabe | Konzentration an gebildetem NADH in µmol/ml |
| Nr.1 | 0,255 | 0,0410 |
| Nr.2 | 0,586 | 0,0942 |
| Nr.3 | 0,912 | 0,147 |
In der Küvette lagen folgende Ausgangskonzentrationen
vor der Phosphatzugabe vor (die Angabe der Fructose 1,6 bisphosphat-Anfangskonzentration
bezieht sich auf die Stoffmenge, die ohne die Anwesenheit der Aldolase
in dem Volumen erreicht würde):
| Phosphatzugabe | Anfangskonzentration Fructose 1,6 bisphosphat in µmol/ ml | Anfangskonzentration an Phosphat (gesamt) in µmol/ml | Anfangskonzentration an NAD+ in µmol/ml |
| Nr.1 | 10,111 | 0,505 | 5,050 |
| Nr.2 | 9,910 | 2,47 | 4,950 |
| Nr.3 | 9,718 | 6,31 | 4,854 |
Die Konzentration an H+ in der Küvette ist 1.10-8 mol/l = 1.10-5 µmol/ml.
Über die Konzentration von NADH im Gleichgewicht können die Konzentrationen von Phosphat und NAD+ im Gleichgewicht bestimmt werden.
{NAD+}Ggw={NAD+}0-{NADH}
{Pi}Ggw={ Pi }0-{NADH}
Über den Wert für DG0I= 24kJ/mol läßt sich die Gleichgewichtskonstante der Aldolase-Reaktion berechnen.
KíAldolase=6,18.10-2 µmol/ml.
Mit diesen Werten läßt sich nun die "chemische" Gleichgewichtskonstante für die Glycerinaldehyd-3-phosphat-Dehydrogenase berechnen.
Hierzu berechnet man die Gleichgewichtskonzentrationen an DHAP und GAP über die Gleichungen:
![]()
{GAP}={DHAP}-x
x={NADH}
Diese Konzentration setzt man dann in Gleichung 13 der Versuchsbeschreibung ein:
![]()
Man erhält die "chemische Gleichgewichtskonstante".
Man berechnet diese Konstante anstatt mit einer Protonenkonzentration
von 1.10-5µmol/ml mit 1.10-1µmol/ml.
(Statt beim pH-Wert 7 (1.10-7mol/l), der Bedingung
für die "biochemische Gleichgewichtskonstante", wurde der Versuch
mit pH-Wert 8 (1.10-8mol/l) durchgeführt; das
entspricht einer Abweichung um den Faktor 0,1.)
| Phosphatzugabe | Gleichgewichts-konzentration von GAP in µmol/ml | KGAPDH | KíGAPDH | DGoë in kJ/mol |
| Nr.1 | 0,7599 | 9,51.10-9 | 0,0951 | 5,83 |
| Nr.2 | 0,7062 | 1,089.10-8 | 0,1089 | 5,49 |
| Nr.3 | 0,6295 | 1,177.10-8 | 0,1177 | 5,30 |
| Mittelwerte | 0,6985 | 1,073.10-8 | 0,1072 | 5,54 |
(Antwort für Aufgabe 4:)
Damit folgt nach dem Heßëschen Satz für die Hydrolyse des Carbonsäure-Phorsphorsäure-Anhydrids im 1,3-Bisphosphoglycerat ein D G0ë von 48,64 kJ/mol.
Nun kann man mit der berechneten Gleichgewichtskonstanten für die GAPDH die Gleichgewichtskonzentration von GAP nach der Zugabe der Triosephosphatisomerase berechnen.
DExtinktion bei dieser Reaktion = 0,84
Das entspricht einer Konzentration von NADH: 0,1350µmol/ml
Über die Gleichung
![]()
erhält man die Gleichgewichtskonzentration von GAP.
Die Gleichgewichtskonzentration des DHAP erhält man durch das Aldolase-Gleichgewicht, über die Gleichung:
![]()
Mit einer Phosphatkonzentration von 6,28µmol/ml,
einer Fructose 1,6 Bisphosphatkonzentration von 9,66µmol/ml
und der Konzentration von NAD+ 4,83µmol/ml
erhält man für den Mittelwert der KGAPDH die Werte:
{GAP}=0,5907µmol/ml
{DHAP}=1,081µmol/ml
Setzt man diese Konzentrationen zueinander ins Verhältnis, erhält man die Gleichgewichtskonstante der Triosephosphatisomerase.
![]()
Daraus folgt für die Reaktion der Triosephosphatisomerase ein DG0ë-Wert von 1,496 kJ/Mol.
Der Literaturwert für diese Reaktion ist: DG0ë=7,55KJ/mol.
Damit weicht unser Wert deutlich von dem Literaturwert ab. Daher ist ein systematischer Fehler oder ein Fehler in der Auswertung leider nicht auszuschließen.
Aufgabe 6:
Siehe Anhang
Aufgabe 7:
In diesem Diagramm ist die Extinktion von NADH bei
340 nm, die bei den beiden Versuchen gemessen wurde, als gekoppelter optischer
Test gegen die Zeit aufgetragen.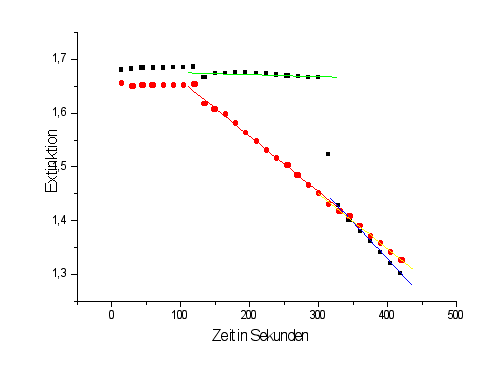
Beschriftung im Diagramm:
Quadrate: Versuch 1
Kugeln: Versuch 2
Aufgabe 8:
Die Steigung dem Graphen Funktion kann dann über den Extinktionkoeffizienten in die Reaktionsgeschwindigkeit der Reaktion umgerechnet werden.
Aus der Reaktionsgeschwindigkeit pro Sekunde ergibt sich die Volumenaktivität µmol/ml.min, indem man die Geschwindigkeit pro Minute ausrechnet.
Dieser Wert gibt die Aktivität der Enzymmenge in der Küvette an. Da in dem Versuch 5 µl Aldolase Lösung eingesetz wurden, berechnet man nun die Aktivität, die 1 ml Enzymlösung hat, aus. Diesen Wert muß man dann noch durch zwei teilen, da die Aldolase nur halb so schnell ist wie die Reaktion der Glycerinphosphatdehydrogenase. Die Ursache hierfür ist, daß die Aldolase pro Zeiteinheit ein Molekül Fructose 1,6 Bisphosphat zu einem Molekül Dihydroxyacetonphosphat und einem Molekül Glycerinaldehydphosphat umsetzt. Glycerinaldehydphosphat wird über die Triosephosphatisomerase in Dihydroxyacetonphosphat umgesetzt. Diese Reaktion ist so schnell, daß man davon ausgehen kann, daß auch alles Glycerinaldehydphosphat verbraucht wird.
Aus dem Diagramm wurden folgende Steigungen berechnet und in eine Aktivität umgerechnet:
Steigung der Funktion:
(Y) = A + B x
Parameter Value Error
------------------------------------------------------------
A 1,67891 0,0042
B -3,63636E-5 1,87781E-5
------------------------------------------------------------
d{Aldolase}/dt=2,92.10-5µmol/ml.s
Enzymaktivität in der Küvette :![]() =1,75.10-4
µmol/ml.min
=1,75.10-4
µmol/ml.min
Enzymaktivität der Stammlösung: 4,87.10-3![]() .
.
(Y) = A + Bx
Parameter Value Error
------------------------------------------------------------
A 1,76491 0,0042
B -0,00104 1,87781E-5
------------------------------------------------------------
Reaktionsgeschwindigkeit für die Aldolase Reaktion:
![]()
d{Aldolase}/dt=8,36.10-5µmol/ml.s
Enzymaktivität in der Küvette :![]() =5,016.10-3µmol/ml.min
=5,016.10-3µmol/ml.min
Enzymaktivität der Stammlösung: 5,016.10-3![]() .
.
3. Die Steigung im Versuch 1 nach der Zugabe der unverdünnten Enzymlösung:
Steigung der Funktion:
(Y) = A + Bx
Parameter Value Error
------------------------------------------------------------
A 2,04625 0,09385
B -0,00181 2,54255E-4
------------------------------------------------------------
d{Aldolase}/dt=1,45.10-4µmol/ml.s
Enzymaktivität in der Küvette :![]() =8,73.10-3µmol/ml.min
=8,73.10-3µmol/ml.min
Enzymaktivität der Stammlösung: 8,73.10-3![]() .
.
4. Die Steigung im 2. Versuch nach der 2. Zugabe an unverdünntem Enzym:
Steigung der Funktion:
(Y) = A + Bx
Parameter Value Error
------------------------------------------------------------
A 1,75275 0,01011
B -0,00101 2,73896E-5
------------------------------------------------------------
d{Aldolase}/dt=8,12.10-5µmol/ml.s
Enzymaktivität in der Küvette :![]() =4,87.10-3µmol/ml.min
=4,87.10-3µmol/ml.min
Enzymaktivität der Stammlösung: 4,87.10-3![]() .
.
Aufgabe 9:
In diesem Versuch wird deutlich, daß bei einem gekoppeltem optischen Test die Bedingungen so gewählt werden müssen, daß die nachgeschlatete Reaktion wesentlich schneller abläuft als die Reaktion, die in dem Test untersucht werden soll.
Im ersten Versuch ließ sich die Reaktionsgeschwindigkeit durch weitere Zugabe von Glycerin-3-phosphatdehydrogenase weiter steigern. Das heißt, daß die Reaktion der Aldolase im ersten Versuchsteil schneller war als die der Glycerin-3-phasphatdehydrogenase und mehr Produkte für die Glycerin-3-phasphatdehydrogenase gebildet hat, als diese verbrauchen konnte. Unter diesen Bedingungen kann die Geschwindigkeit und damit die Enzymaktivität nicht richtig bestimmt werden.
Wie sich jedoch im zweiten Versuch zeigt, ist die zweite Zugabe von Enzym im ersten Versuch ausreichend, um die Geschwindigkeit der Glycerin-3-phosphatdehydrogenase größer als die der Aldolase werden zulassen. Damit kann aus Versuch 1 mit Hilfe von Versuch 2 auch die Geschwindigkeit der Aldolase bestimmt werden.
Im Versuch 2 zeigt sich, daß sich durch die Zugabe von weiteren 5µl unverdünnter Enzymlösung die Geschwindigkeit der Reaktion nicht weiter gesteigert werden kann. Das bedeutet, daß die Aldolase-Reaktion schon im ersten Versuchteil von Versuch 2 langsamer als die Reaktion der Glycerin-3-phosphatdehydrogenase war. Damit kann unter diesen Bedingungen die Aktivität der Aldolase richtig bestimmt werden.
Zu bemerken ist, daß die Reaktionsgeschwindigkeit der Aldolase-Reaktion im ersten Versuch nach der zweiten Zugabe des Enzyms größer ist, als die Geschwindigkeiten in Versuch 2.
Für diese Beobachtung habe ich keine theoretische Erklärung. Es sollte daher ein Pipettierfehler vorliegen.